 Luke
Barrett
Luke
Barrett
CPBR Summer Scholarship
Supervisor: Peter Thrall
The study of fungal diseases in natural plant communities can contribute significantly towards the understanding of evolutionary and ecological dynamics that occur within plant host-pathogen systems. In this study, I have investigated how variation in aspects of growth of the fungal pathogen, Alternaria brassicicola may provide insight into the evolution and ecology of that species. This project is part of a broader study aiming to address questions regarding the role of population and spatial structure on disease dynamics within the Cakile maritima-Alternaria brassicicola host-pathogen interaction on the southern coast of NSW. Because Cakile only grows on beach foredunes and strandlines, it is easy to delineate populations, making this system ideal for spatial studies. The study area includes 60 populations in 3 subregions along the south coast of NSW. As part of the broader study, a large collection of pathogen isolates was generated to examine spatial structure between populations. These isolates are stored in silica gel and are easy to re-culture for experimental purposes. While isolates were being cultured, it was noticed that there was considerable variation in growth rates. This raised the question of whether isolates which varied in growth rate in culture also differed in their ability to infect plants, the severity of disease caused, and in the amount and rate of spore production.
Equally, it is of interest to ask by what means this variation is being maintained within the broader population. If, for example, higher growth rate leads to a competitive advantage, with fast growing isolates able to more effectively colonise and utilise host tissue than slow growing isolates, then directional selection for faster growing isolates may be expected. Trade-offs between components of fitness may be one means by which diversity is maintained within natural populations. Genetic trade-offs could maintain polymorphisms for pathogen aggressiveness within pathogen populations, since genetic gains associated with one fitness component, may be offset by genetic losses associated with another fitness component, resulting in equal fitness overall. For example, while aggressive pathotypes may be able to more effectively compete for limiting resources, they may be less efficient at producing spores than less aggressive pathotypes. Selection may then favour each pathotype equally, but for different reasons. Genetic costs of increased aggressiveness may be demonstrated in this case through a significant negative correlation between growth rate in culture and spore production in culture, providing that a positive correlation can be shown between growth rate in culture and aggressiveness on the plant.
To explore these possibilities a study was designed to investigate variation in growth and spore production in the Alternaria isolates. The data collected were used to address the following questions:
The study involved both laboratory and glasshouse components. The laboratory component involved culturing a range of the available isolates and characterising them for growth. Alternaria brassicicola isolates were cultured on Potato Dextrose Agar (PDA). All isolates were grown under identical conditions for 10 days in a 25oC growth chamber. Colony diameters were then measured along two perpendicular lines. The distribution of growth rates within the entire set of isolates was determined. Considerable variation in growth rates was observed, with significant among-population differences.
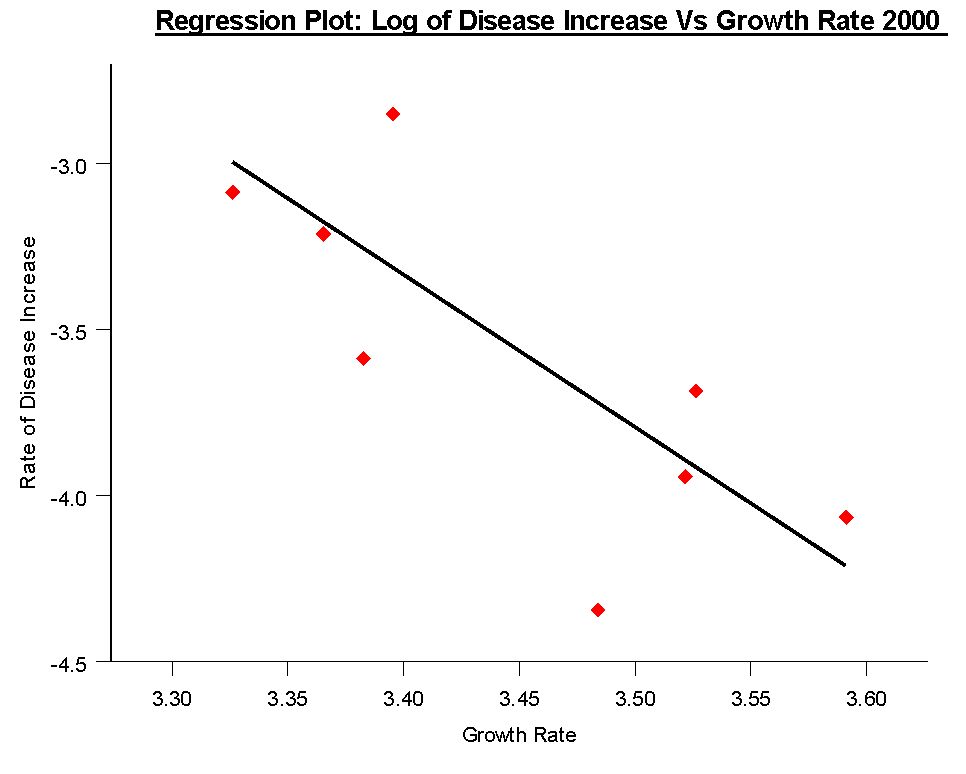
Because disease epidemic development was censused at regular intervals in the natural populations from which these isolates were collected, I was able to compare the mean growth rate of isolates in culture for separate populations, and the rate of epidemic development within those populations. A significant negative relationship between epidemic development and mean population growth rates was evident.
A subset of 20 isolates, ten fast growing and ten slow growing, were then selected from each end of the distribution in order to establish that fast and slow growing isolates were significantly different from each other, and for use in assessing spore production and performance on Cakile plants. Each treatment was replicated ten times and grown together in a randomised block design under identical conditions for 10 days in a 25oC growth chamber. There was a highly significant difference (P < 0.01) between the fast and slow growing groups.
To assess spore production, conidial counts were made for 5 replicates within each treatment group. Conidial suspensions were prepared from 10 day old cultures by adding 2.5 ml of 70% ethanol to each plate and rubbing the surface of each culture with a spreader bar. Conidia were counted with a hemacytometer. A negative relationship between growth rate and spore production per unit area was found.
 Glasshouse
Trials
Glasshouse
Trials In order to assess whether or not growth rate in culture is indicative of aggressiveness on the plant, a glasshouse inoculation trial was performed. Each of the subset of twenty isolates was seperately inoculated onto a set of 15 Cakile plants in order to assess isolate aggressiveness. Cakile plants were grown from randomly selected and mixed seed. Each isolate suspension was adjusted to a standard concentration using a hemacytometer. Spore suspensions were then atomised onto each treatment group. Each treatment group was covered with a tent of clear plastic, preventing cross-contamination and raising humidity levels. The aggressiveness of each isolate was assessed by scoring each plant in the treatment group for percent of diseased tissue. The results from this trial have yet to be analysed.
There is a high degree of variability in growth rate among Alternaria brassicicola isolates. This variability appears to be under genetic control. In addition to differences between individual isolates, there are also significant differences between pathogen populations in terms of their mean growth rates. There is a negative relationship between mean population growth rate and calculated rate of disease prevalence increase. A negative relationship between spore production per unit area and growth rate was also found. Viewed together, these two negative relationships provide good circumstantial evidence for a trade-off between growth rate and spore production. Slower growing isolates, while perhaps not able to compete in terms of growth, may be able to produce spores and hence colonise new plants more effectively than faster growing isolates.