![An Australian Government Initiative [logo]](/images/austgovt_brown_90px.gif)

Mistletoes - evolution
Floral evolution: embryology and fruit structure
Mistletoes are so unusual in the structure of their floral ovaries that one student of the families, van Tieghem, was tempted to treat them as a separate class of flowering plants, with status equal to the monocotyledons and dicotyledons. However these unusual features form a striking sequence which shows how they were derived by progressive reduction from the typical ovary structure of flowering plants. The special features of ovary structure and embryology may be related to the adaptations in mistletoes for aerial parasitism.
The modifications of ovary structure and embryology reach their extreme in the southeast Asian genus Helixanthera ![]() . Like most flowering plants, mistletoes have a female reproductive system comprising an ovary, at the base of the flower, with a columnar style arising from it, and a receptive stigma (for pollen) at the apex of the style. But unlike normal flowering plants, in Helixanthera there are no chambers in the ovary, bearing the ovules which become the seeds after fertilization. In Helixanthera the ovary is solid, and there is a small mass of reproductive tissue at the base of the canal which runs down the centre of the style into the ovary.
. Like most flowering plants, mistletoes have a female reproductive system comprising an ovary, at the base of the flower, with a columnar style arising from it, and a receptive stigma (for pollen) at the apex of the style. But unlike normal flowering plants, in Helixanthera there are no chambers in the ovary, bearing the ovules which become the seeds after fertilization. In Helixanthera the ovary is solid, and there is a small mass of reproductive tissue at the base of the canal which runs down the centre of the style into the ovary.
In normal flowering plants a reproductive cell in the ovule divides to produce an embryo sac, which contains an egg cell. Pollen grains germinate on the stigma, and produce filamentous pollen tubes which grow down the stylar canal to the ovule, where a male cell from a pollen tube fertilizes the egg, and the fertilized ovule begins its development into a seed, within the ovary as it developes into the fruit. However in Helixanthera several reproductive cells in the solid ovary divide to produce embryo sacs, which then elongate, growing up the stylar canal, carrying eggs right to the stigma. The pollen tubes therefore only grow a very short distance to meet the eggs.
In Helixanthera the fertilized eggs therefore start their development into embryos not in the ovary but at the base of the stigma. The embryos rapidly produce a filament of cells (the suspensor) which pushes the successful embryo back down the stylar canal into the ovary before the style withers and falls off! Thus the embryo is in place inside the fruit as it developes its nutritive and sticky layers. So Helixanthera is remarkable in not having ovules (and therefore true seeds) and in sending its eggs to meet the pollen instead of vice versa.
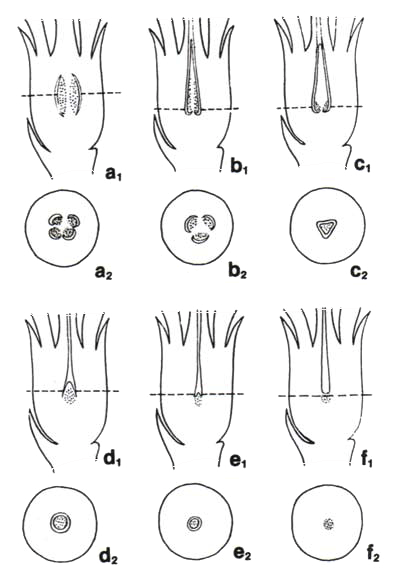
The evolutionary sequence leading to the situation in Helixanthera is shown by the situation in some other genera (below). In the Australian genus Lysiana (a1, a2) ![]() the ovary actually has four chambers. In each of these chambers there is a lobe of tissue which probably represents a much-reduced ovule. Embryo sacs are produced in each of these lobes, and do elongate, but only reach partway up the stylar canal. Thus the pollen tubes grow much of the way to meet the eggs. In the southeast Asian genus Lepeostegeres (b1, b2) there are only partial partitions between the chambers, and there is a central pyramidal mound of tissue, three-lobed at the base, with embryo sacs produced in the lobes. The pyramidal mound extends partway up the stylar canal, but the embryo sacs do not extend any further. In the southeast Asian genus Macrosolen (c1, c2)
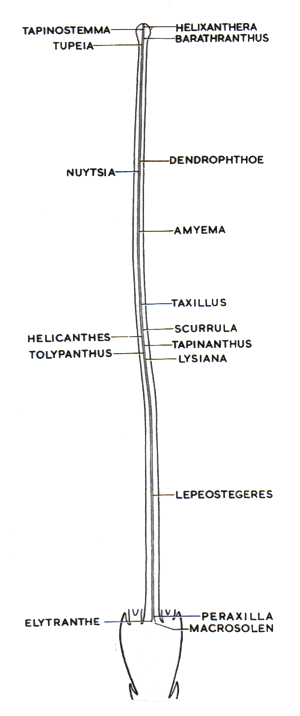
the ovary actually has four chambers. In each of these chambers there is a lobe of tissue which probably represents a much-reduced ovule. Embryo sacs are produced in each of these lobes, and do elongate, but only reach partway up the stylar canal. Thus the pollen tubes grow much of the way to meet the eggs. In the southeast Asian genus Lepeostegeres (b1, b2) there are only partial partitions between the chambers, and there is a central pyramidal mound of tissue, three-lobed at the base, with embryo sacs produced in the lobes. The pyramidal mound extends partway up the stylar canal, but the embryo sacs do not extend any further. In the southeast Asian genus Macrosolen (c1, c2) ![]() there are no partitions forming chambers, so that the ovary has a single cavity. Again there is a pyramidal mound of tissue in the cavity, three-lobed at the base, with embryo sacs produced in the lobes. In other genera in the sequence the pyramidal mass is smaller and smaller (d1, e1, f1), the lobes less and less developed, and the cavity smaller and smaller (d2, e2, f2) , until we reach the state in Helixanthera with no cavity, no pyramidal mass, no lobes and just a single mass of reproductive tissue at the base of the stylar canal (f2). In these various intermediate states the embryo sacs extend to various heights in the stylar canal (below right). The largest Australian genus, Amyema, is relatively advanced in the sequence, similar to Helixanthera except that the embryo sacs only reach two-thirds the way up the style.
there are no partitions forming chambers, so that the ovary has a single cavity. Again there is a pyramidal mound of tissue in the cavity, three-lobed at the base, with embryo sacs produced in the lobes. In other genera in the sequence the pyramidal mass is smaller and smaller (d1, e1, f1), the lobes less and less developed, and the cavity smaller and smaller (d2, e2, f2) , until we reach the state in Helixanthera with no cavity, no pyramidal mass, no lobes and just a single mass of reproductive tissue at the base of the stylar canal (f2). In these various intermediate states the embryo sacs extend to various heights in the stylar canal (below right). The largest Australian genus, Amyema, is relatively advanced in the sequence, similar to Helixanthera except that the embryo sacs only reach two-thirds the way up the style.
 |
 |
Thus in mistletoes we find progressive reduction in the normal structure of the ovary, to a point where there is no true seed formed within the fruit, and instead a well-developed embryo lies within the solid tissues of the fruit itself. This is presumably yet another aspect of the adoption of the mistletoe lifestyle.