 |
Centre for Australian National Biodiversity
Research
|
CPBR NEWS
Issue 59 - March 2004
News from the Centre for Plant Biodiversity Research, Australian National Herbarium (CANB), for the information of CPBR and ANBG staff and volunteers.
CPBR News is produced monthly. If you wish to contribute, please email your suggestions to Val Oliver, the coordinator.
Val Oliver: ph (02) 6246 5533; fax (02) 6246 5249; email: Val.Oliver@csiro.au
1. Staffing News
The Centre welcomes Luke Bulkeley who commenced full time this week. Luke is working half time on the Healthy Country project and half time in subprogram UB managing the germplasm collections
[Val Oliver]
********************
2. Australian National Herbarium
Student Botanical Internship Program 2004 Overview
Participants and Institutions
2004 represents the twelfth year that the Student Botanical Intern Program has been run. A total of 19 applications were received for the Program, with 18 placements offered and 17 accepted. All 17 Interns successfully completed the Program in 2004. The number of applications received this year was comparable to numbers in recent years.
Recruitment Trends
Nine Australian universities or colleges were represented along with graduates from an overseas institution. There were five Australian National University, two Canberra University and two CIT students making the 2004 Program a Canberra dominated one. The trend of low numbers from Sydney continues, with only one student taking part in the program. A student from Germany also helped to provide an international flavor to the Program.
Many of our participants had completed, or were in the process of completing, degrees in horticulture. Other students were working towards degrees in environmental science, botany or geography. Overall it was a good mix of backgrounds and interests that no doubt added to the Intern’s learning experience.
For the first time a second call for applications was made after disappointing numbers were received at the close of the first application deadline. This may be the result of students needing to find paid work during the summer break to meet university fees. It may also show the direct targeting of academics to sell the program is beginning to fail and future recruitment needs to move to electronic advertisements.
Work Output
Output achieved by Interns during 2004 is roughly equal to 1.4 year’s work (of an entry-level TO), based on a 200-day working year. However, this does not take account of the CPBR and ANBG staff, many of which put in long hours, supervising and lecturing. This comparatively is a reduction on previous years’ productivity (the average over 1999-2003 being 1.7 years). This reduction is largely attributable to the shortening of the Program from eight weeks to seven; forced on us to better fit in with earlier University starting dates.
The major push to process herbarium specimens identified as priority groups for the AVH project continued this year. This work included mounting and incorporation of vascular and non-vascular specimens, identification of specimens, assistance with loans and exchange, data entry, determining specimen geocodes, spirit collection maintenance and general lab and herbarium tasks. The major focus of the curation in 2004 centred around the orchid collection. As part of this additional herbarium space was made available by reshuffling neighbouring families for future incorporation. Significant progress was also made to changing names in the orchid spirit collection.
The unmounting of the remaining Deniliquin Rangeland Herbarium specimens for appropriate archival remounting was a major focus of the Intern curatorial labour. Started in the 2003 Program, this was completed in 2004. Processing of the QRS herbarium’s duplicate backlog was another large task that was continued with the help of this years Interns, good progress being made.
Along with curation work, this year saw a trial of intern research assistance. Each intern was assigned, mostly in pairs, to an ongoing CPBR research project where they provided basic assistance for half a day a week. This work was aimed at providing interns with additional skills and experience along with providing much needed labour to researchers. The biggest project saw all interns over several days recording measurements they took from 500 eucalypt seedlings as part of a CPBR supported PhD project. Assistance was also provided to research groups such as orchids, cryptogams and several ongoing lab projects. The developing "orchid key" interactive product was also tested by several interns who made comment on its accuracy and usability. See Attachment A for detailed work outputs.
After the January 2003 bushfires a new route had to be selected for the annual eucalypt identification fieldtrip. Ian Brooker led the interns through sites around Queanbeyan, Captains Flat and Tallaganda State Forest, introducing them to many of the local tree species. The four-day residential field trip to Jervis Bay went ahead after early fears the trip may have to be cancelled due to bushfires in the area. Much of Booderee National Park was burnt, but this allowed the opportunity for Malcolm Gill to lead the interns in a post fire assessment of the woodland and heath communities. Species showing varying degrees of response to fire were collected, identified and documented. The students also enjoyed a bush tucker tour led by Wreck Bay local Barry Moore around the Booderee Botanic Gardens.
Publicity
The 2004 Program attracted media attention at the end of the course with an ABC 666 Local Radio interview of two graduates and a short Canberra Times article. There was also a full-page article published in the Summer 2004 Natural Heritage detailing the activities of the 2003 Intern program.
Evaluation
Overall, the 2004 Interns Program was very successful. Work output was good if somewhat briefer in duration than previous years. A wide range of curatorial and research tasks were supplemented with much needed intern labour. The general bulk processing of herbarium specimens still makes up the core component of intern work each year, work that is sorely needed to keep the collection up to date.
All Interns were also provided with Evaluation Forms for the Program. Comments from these forms are currently being reviewed to help make improvements to the 2005 Program.
Budgeting
Overall costs come out at $5666 to run the 2004 Interns Program, slightly more expensive than 2003 (at $5450). As with previous years most of this cost was generated by the Jervis Bay fieldtrip, and associated vehicle hire and fuel costs.
Concerns and Issues
No major difficulties were encountered during the running of the 2004 Program, though a few issues need to be thought about for next year:
Several key institutions (eg. ANU and Uni of Canberra) began their first semesters in the last week of February, resulting in the traditional eight-week program for the first time being reduced to seven weeks. It seems likely that this will have to be a permanent change, resulting in a reduced output. The recommendation is that several non-core lectures are cut from the program and perhaps more students be recruited to help reduce this loss of productivity.
IT support was much improved as compared to previous years, the in-house presence of Kim Navin provided quick and effective support when computer problems were inevitably encountered. Email was once again set up on the EA network and linked to Intern personal web accessible accounts in order for them to be to readable on the CSIRO network; whilst this works a simpler solution would be preferred.
The perennial problem of accommodation for interstate participants remains a major issue. All of the western edge ANU colleges (eg. Burgmann, Burton and Garran) were fully booked before interns could even apply for residency. This year a deal was done with Fenner Hall to pre-book places for interns for the full seven weeks; several students took up this offer. This seems to be the way to go with future programs, a deal with Fenner or possibly a closer college will help provide some assurance to incoming students in Canberra’s ever tightening rental market.
Acknowledgements
The success of the 2004 Program is in no small part due to the considerable efforts of a number of Centre staff, outside academics and others who freely gave their time to present lectures and training sessions, as well as providing supervision for Intern work teams. I am most grateful to all concerned. Thanks are also due to all CPBR and ANBG staff, especially those at the Herbarium, for their tolerance, enthusiasm and support during the course of the Program.
[Anthony Whalen]
********************
3. Research Groups
Research Reports from Summer Students 2003-2004
Phylogenetic relationships of the Australasian citrus: the Citreae tribe in a world context.
Sarah Rich Supervised by Randy Bayer
The agricultural production of citrus has flourished worldwide for centuries due to both its ornamental and culinary value. The citrus fruits and their wild relatives belong to the Rutaceae family, within the subfamily Aurantioideae. The subtribe Citrineae (tribe Citreae) is a group of these plants which are unique in bearing fruits containing pulp vesicles that fill all the space in the segments of the fruit not occupied by the seeds. They are almost exclusively native to China and the South Pacific region, including Australia. Their unique fruit are the basis for one of the most important fruit industry in warm countries of the world due to the great commercial importance of the juice extracted from species of the genus Citrus (Mabberley, 1998; Swingle and Reece, 1967).
Australia is the fourth largest citrus producing country in the Southern Hemisphere, with the citrus industry having an estimated gross production value of around $4.3 million (Anon, 2004). Wild members of Australasian Citreae are anticipated to possess genes which provide a number of beneficial traits for growth in the Australian environment and therefore it is predicted they will be useful in citrus breeding programs. Breeding programs are already investigating native members resistance to salt, boron and the cold but the breeding of new, high quality Citrus cultivars for the Australian environment is dependent on reliable information about the relationships of species in genera within the tribe Citreae (Birmingham, 1998).
Over the last 250 years a number of taxonomic classifications have been proposed for the plants contained within the tribes and subtribes of the Aurantioideae. The most recent is that of Swingle and Reece (1967) although Tanaka’s original treatment (1936) and more recent classifications are often referred to. The tribe Citreae contains three subtribes the Citrineae, Triphasiinae and the Balsamocitrinae. Within the Citrineae the genus Citrus itself has been variously described as consisting of from 1 to 162 species. Swingle’s widely accepted 1946 classification system recognises 16 species while Tanaka (1977) recognises 147 species (Moore, 2001). Closely related to the genus Citrus are several genera, which were recognized by Swingle, including the genera Clymenia, Eremocitrus, Fortunella, Microcitrus, and Poncirus. Recent work has questioned the distinctness of these genera and some botanists now include them within an enlarged concept of the genus Citrus (Mabberley, 1998).
Relationships among species and genera of the group are complicated by its long history of cultivation as well as a high frequency of bud mutation, a wide cross-compatibility among species as well as many species having some degree of apomixis (asexual reproduction in which plant embryos grow from egg cells without fertilisation by pollen), which tends to reduce variability within species leading to a proliferation of named species (Moore, 2001). CSIRO is aiming to provide a molecular-based evolutionary tree of relationships of members within the tribe Citreae based on chloroplast DNA sequences. The summer project concentrated on 16 species of citrus, mostly Australasian, sequences from these were to be combined with those already elucidated in earlier research.
DNA was extracted from the leaves of fresh frozen samples from the US Department of Agriculture and dry samples from CSIRO’s Merbein Research Station. The polymerase chain reaction (PCR) was used to amplify three regions from the chloroplast genome; the trnL intron and trnL - trnF intergenic spacer (1300 base pairs); the rpsL16 intron (1460 base pairs) and the atpB-rbcL intergenic spacer region (1100 base pairs) of the chloroplast genome. The amplified PCR product was then cleaned and used to obtain sequences. These were manually aligned, with sequences downloaded from Genbank, for use in cladistic analysis. Phylogenetic analysis was carried out in PAUP* 4.0, employing a maximum parsimony, heuristic search with uninformative characters left out. Searches were conducted using a tree bisection-reconnection (TBR) branch swapping algorithm with all possible shortest trees saved to a maximum of 100000; from these, both strict and 50% majority consensus trees were computed. The time restraints of the summer scholarship program did not allow for a full analysis to be completed, however the preliminary findings were most encouraging.
Figure One: Strict consensus tree of combined rps16 intron and the trnL intron and spacer regions.n = 40; Informative characters = 201;
Shortest tree length = 393; Rohlf’s CI = 0.638; Outgroup = 2 (Ruta graveolens, Chloroxylon swietenia). * = sequenced by S.Rich
In essence all the trees produced support Swingle’s treatment by both the monophyletic nature of genera and by the presence of subtribal groups from his treatment, although with the limited number of taxa studied it is difficult to draw definitive conclusions. Figure one shows a strict tree of the combined rps16 intron and the trnL intron and spacer regions. This tree illustrates all of the trends found across the five consensus trees produced and highlights our most important finding; that all the Australasian members of the Citrinae subtribe appear to of monophyletic lineage, as suggested by Mabberley’s (1998) treatment of these taxa. Also falling within this clade in all trees were both Oxanthera neocaledonica and Clymenia polyandra, suggesting that they are both possible members of the Citrus genus. Given their origins (New Caledonia and Papua New Guinea, respectively) as well as their taxonomic histories this was not a particularly surprising find, but one that would definitely bear further investigation. Another surprising result concerned the genus Atalantia, which unlike all other genera studied does not appear to be monophyletic. In all trees, apart from that of the trnL intron and spacer region, Severinia buxifolia and Atalantia ceylanica appeared as a monophyletic clade. Originally part of the Atalantia genus, five of the six Severinia species were only segregated from Atalantia in 1938 by Swingle, although they are morphologically very similar to Atalantia. With sequences from only four of the eleven Atalantia species and one Severenia, our results merely highlight the need for further work on these genera.
Overall the results produced at the end of the ten week summer program raised more questions than they solved, however as a preliminary study into the Australasian citrus, this project was successful in highlighting the areas of dissonance within the current classification system. Hopefully with additional work and a more in depth analysis of the data already collected it will be possible to further elucidate the origins and relationships found within the tribe Citreae.
Acknowledgements
Thanks to The Centre for Plant Biodiversity Research and CSIRO for providing me with the Summer Scholarship and the opportunity to work on this project. Special thanks to Randy Bayer for all the enthusiasm and encouragement during his supervision, along with the entire molecular systematics lab group for their patience and support
References
Anonymous. 2004 (last updated). Australian Citrus Growers Inc.[ONLINE] http://www.austcitrus.org.au/internal.php?page_id=6
Accessed: 13 February 2004
Birmingham, E. 1998. ‘Australian native citrus: wild limes from the rainforest to the desert…’ The Australian New Crops Newsletter Issue 10.
[ONLINE] http://www.newcrops.uq.edu.au/newslett/ncn10211.htmAccessed:
29 January 2004
Mabberley, D J. 1998. ‘Australian Citreae with notes on other Aurantioideae (Rutaceae).’ Telopea Vol 7(4): 333 – 344.
Moore, G A. 2001. ‘Oranges and Lemons: clues to the taxonomy of citrus from molecular markers.’ Trends in Genetics Vol 17(9): 536 – 540.
Swingle, W. T. and Reece P. C. 1967. The botany of Citrus and its wild relatives. IN Reuther W., Webber H. and Batchelor L. [eds.] The Citrus Industry,
Vol 1.p: 190 – 430.University of California, USA.
Conservation of the endangered Australian terrestrial orchid, Oligochaetochilus gibbosus, through symbiotic seed germination, and the use of seed kept in long-term storage.
Emma Crewdson
Supervisor: Mark Clements
 Oligochaetochilus
gibbosus, the Illawarra Greenhood Orchid, is an endangered orchid species
that is endemic to the NSW central coast. Although protecting the habitat
of this species is an important conservation strategy, artificial propagation
is another method that could be used to help conserve O. gibbosus.
This would involve germinating seed collected from wild plants, and then the
translocation of seedlings back into their natural habitat.
Oligochaetochilus
gibbosus, the Illawarra Greenhood Orchid, is an endangered orchid species
that is endemic to the NSW central coast. Although protecting the habitat
of this species is an important conservation strategy, artificial propagation
is another method that could be used to help conserve O. gibbosus.
This would involve germinating seed collected from wild plants, and then the
translocation of seedlings back into their natural habitat.
In the wild, all orchids require a relationship with a mycorrhizal fungus in order for their seed to germinate. This relationship is often used when artificially propagating orchids from seed (the technique is known as symbiotic seed germination) in order to create seedlings with a higher chance of survival than those grown through asymbiotic methods.
When thinking about how to do this, several questions arise. If the seed is to be symbiotically germinated, which fungus will be most successful in terms of germination rate? Also, there is some O. gibbosus seed that has been in long-term storage for up to 17 years, including some that was collected from an area where O. gibbosus is now virtually extinct. It is important that the viability of this seed, and hence its potential to contribute to the conservation of O. gibbosus is determined, particularly with relation to preserving the genetic diversity of the species, a vital factor when attempting to conserve this orchid.
Hypotheses
These questions lead to two hypotheses that were tested in several experiments
1. There are no differences between the effects of different mycorrhizal fungal isolates in symbiotic seed germination of O. gibbosus?
2. There are no differences in seed germination of O. gibbosus when using stored seed of varying ages.
Experiment 1
The purpose of this experiment was to determine the effectiveness of various mycorrhizal fungi. Hence, 6 fungal isolates were chosen: 2 from O. gibbosus, 2 from Hymenochilus bicolor (a closely related genus to Oligochaetochilus, collected from sites where O. gibbosus was present), 1 from O. rufus (collected from sites where O. gibbosus was not present) and 1 from O. leptochilus (an orchid from Western Australia).
Experiment 2
For this experiment, the variable was seed, with seeds stored for different amounts of time being used. The seed was originally collected in: 1997 (stored at room temperature), 1985, 1982, 1981, 1980, 1978 and 1976.
Methods
For both experiments the method was basically the same:
- Petri dishes containing oat meal agar were inoculated with the appropriate fungal isolate. Oat meal agar was used to ensure that any germination was due to the presence of the mycorrhizal fungi, as opposed to some other media which may allow the orchid seed to germinate without the fungus.
- Next, the seeds to be used were surface sterilised using bleach and then rinsed. They were then sown onto the agar plates. The plates were sealed with Parafilm to prevent contamination.
- Approximately 6 weeks after sowing, the plates were assessed and data collected on the germination stage of the seeds present.
Data Collection
Data was taken by scoring seeds present in a small sample area according to the germination stage they were at, using a scale of 0 to 6 (where 0 is dry seed, no germination, and 6 is a seedling ready to be transferred to soil). Due to the short period that the experiments had been running for, the most developed protocorms had only reached germination stage 2 (breaking the seedcoat).
Results and Discussion – Experiment 1
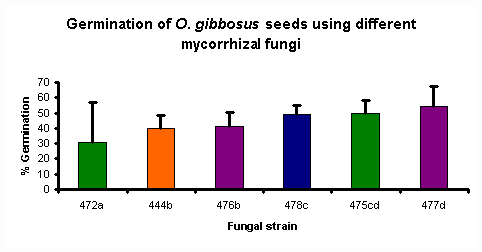
Figure 1
Figure 1 shows the percentage germination of O. gibbosus seeds obtained using the various fungal isolates. Statistical analysis of the results shows that 4 groups of fungi can be formed based on their effectiveness, as shown by the letters a, b, c, d.
It appears that the fungal isolate used will have a definite effect on the percentage germination obtained. However, this effect cannot be linked to species or location of the orchid that the fungus was isolated from. This makes it difficult to predict, when only knowing these factors, the extent to which a particular isolate will enhance orchid seed germination without actually testing it and observing the result.
Results and Discussion – Experiment 2
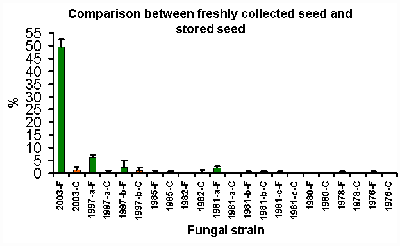 Figure
2
Figure
2
Figure 2 shows the percentage germination obtained using seeds stored for various periods of time. The extremely low germination rates that are present in seed that has been in long term storage (at best, about 6% in comparison to about 50% for the freshly collected seed) indicate that orchid seed does not have the capacity to survive storage in the manner which has occurred in the herbarium. A regression analysis was performed on the data, and the results are shown in figure 3 below.
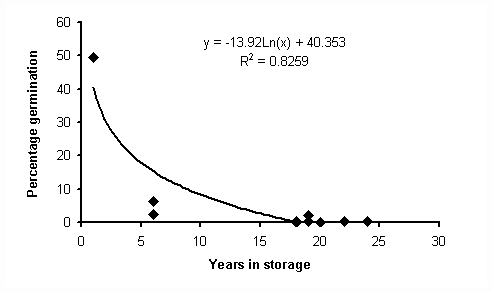 Figure
3
Figure
3
The regression analysis demonstrates that there is an exponential decay in percentage germination over time. However, there is the need for further experimentation to determine the exact shape of the curve (by filling in data gaps, especially between 1 and 5 year old seed).
Conclusions
- there are distinct differences between the effects that different fungal isolates will have on germination percentages when performing symbiotic seed germination, however there appears to be no link between the host species or the location that the fungus is isolated from.
- there is an exponential decay in germination results as older seed is used, however new experiments are required to specify the gradient of this decay curve.
- there may be little point in retaining current seed stores of orchids of this type in the herbarium for an extended period of time, as their viability is unreliable and germination is low.
********************
Patterns of Weed Invasion in Subalpine Vegetation and the Role of Trifolium repens (White Clover)
By Lyndsey Vivian
Supervisor: Bob Godfree
Background
The susceptibility of a vegetation community to invasion is thought to be to a certain extent influenced by the community’s richness (Elton 1958, Naeem et. al. 2000). Indeed many experimental studies have demonstrated a negative relationship between community richness and the richness of exotic species (e.g. Naeem et. al. 2000). One explanation for this is that high native species richness leads to more resources being utilised, leaving fewer resources and niche opportunities for invaders (Shea & Chesson 2002, van Ruijven et. al. 2003). However large-scale observational studies often demonstrate the opposite relationship (e.g. Foster et. al. 2002). These seemingly conflicting results have been explained by the presence of confounding extrinsic factors that are controlled in experimental studies and models (Levine 2000, Shea & Chesson 2002). Shea & Chesson (2002) reconciled these findings by suggesting that the scale of the study will affect the relationship (Figure 1). At a large scale where extrinsic factors vary, a positive relationship may be found, as both native and exotic species richness respond to these factors in the same way. However at a smaller scale where these factors are likely to be more uniform, the number of exotic species is instead reduced by native species richness.
The aim of the study was to investigate patterns of weed invasion in subalpine vegetation both at a large and a small scale, and thus determine whether the richness model of Shea & Chesson (2002) holds true, particularly as the idea that negative relationship even occurs in naturally is highly contentious (Wardle, 2001). We also examined relationships between exotic and native species in terms of biomass, an often neglected means of describing community characteristics.
The second aim of this study was to investigate the distribution and abundance of Trifolium repens, or white clover (Figure 2). T. repens was selected because it is a known invader in subalpine vegetation, and additionally because there are environmental risks associated with the potential release of a genetically modified, virus-resistant T. repens (Godfree et. al. in press).
Methods
The study area, Grassy Creek, is located in Namadgi National Park approximately 65 km south of Canberra, ACT. Grassy Creek lies at 1200 to 1300 metres a.s.l. and is classified as subalpine.
Eight sites were selected from a larger pool of randomly selected sites. These 8 sites were selected to be most representative of the various community types in terms of species composition. Four of the sites were from woodland communities and 4 were from grassland communities.
At each site a 1 m x 1 m quadrat (plot) was laid out, divided into 25 subplots of 20 cm x 20 cm. All above ground biomass (except for large Eucalyptus species) was clipped from each subplot and bagged. Individual species from each subplot were subsequently separated and identified.
All plant material was dried at 95 degrees C for three days until constant weight was achieved and then weighed.
Results and Discussion
Community Characteristics
Grassland communities were dominated by native perennial grasses, such as Poa sp. and Themeda triandra (kangaroo grass). Geranium antrorsum was the most abundant herb. Trifolium repens was the most abundant exotic herb across all grassland sites, although two species of exotic grasses, Poa pratensis (Kentucky bluegrass) and Holcus lanatus (Yorkshire fog) were found in higher abundances.
Across all woodland sites Poa spp. were most abundant. Woodland sites had a shrubby understory consisting of species such as the common Fabaceous shrub Bossiaea foliosa. The overstory was dominated by Eucalyptus stellulata (black sallee). T. repens was relegated to second-most abundant exotic herb in the woodland community behind the Asteracean species Hypochoeris radicata (flatweed), although Holcus lanatus was the most abundant exotic species (and third most abundant species overall in the woodlands).
Richness relationships
At the large scale (across all plots) exotic species richness increased with native species richness as expected. However when this relationship was examined at the small scale (within each plot) there was no evidence of the predicted negative relationship, indeed a positive relationship was evident within some plots particularly in G1 (Figure 3). These relationships were due to the strong positive relationship between native and exotic herbs, the most species-rich functional group in the study area.
The positive relationship between exotic and native herb richness in G1, appears to be related to the observation that both native and exotic herbs decline with increased non-herb biomass (Figure 4). The bulk of non-herb biomass at this high level was due to the presence of a single species not found at any other site, Poa labillardierei (river tussock), a large native tussock grass. This suggests that competition is occurring at a small scale, however is driven by the biomass of a single species, rather than resources being consumed by high richness.
Biomass relationships
Unlike richness relationships, the biomass of native and exotic species were not related at the large scale. However, at the smaller scale, a negative relationship was found in three out of eight plots. Within these plots, it appeared that exotic biomass declined with increasing abundance of the dominant species, such as P. labillardierei, or the dominant functional group, primarily grasses, suggesting competition by a single dominant group/species rather than by high numbers of native species.
We also found that plots containing low amounts of exotic biomass tended to have higher species richness, whereas plots with high amounts of exotic biomass contained few exotic species (Figure 5). It is clear that these two parameters yield different results – many exotic species does not necessarily mean that the biomass of exotics is high. In addition, it appears that where exotic biomass is low (but species-rich) the majority of species are annual/biennual herbs, whereas where exotic biomass is high (but species-poor), all are perennial and the majority are grasses (Figure 5).
|
Av. exotic biomass (g) |
7.0 |
48.3 |
135.7 |
|
Av. biomass of T. repens (g) |
0.51 |
5.87 |
18.44 |
|
Av. exotic richness (no. species) |
6.8 |
6.3 |
3 |
|
Av. exotic species (% total species) |
30.9 |
30.3 |
21.4 |
|
Av. exotic perennial sp. (%) |
39.2 |
80.0 |
100.0 |
|
Av. exotic herbaceous sp. (%) |
87.2 |
77.3 |
33.3 |
Trifolium repens
T. repens was found to co-invade with other exotic species (Figure 6), indicating that T. repens is unlikely to be found in high abundances on its own; it occurs where other exotics also perform well. The relationship between T. repens and native grasses was also examined. In only one instance was there evidence that T. repens was limited by grass biomass, possibly because only once did grass biomass reach this magnitude (driven by the presence of Poa labillaridierei). This indicates that T. repens will not be limited by native grass biomass unless it is very high, which is rare in the communities studied and requires the presence of large tussock grass species such as P. labillardierei.
Conclusions
Measurements of biomass and richness are two different ways of describing a community and do not necessarily follow the same relationships. Our data suggests that when investigating invasions we must ask whether it is the richness or biomass of exotics that has the greatest effect on communities, since models such as that postulated by Shea & Chesson (2002) do not take into the biomass of exotic species. In reality, however, many invaded vegetation communities are overrun by a dominant, vigorous exotic invader.
At the small scale there appears to be a competitive process whereby a single dominant species, or a dominant functional group is able to limit other species. Therefore at the small scale, relationships in subalpine vegetation do not seem to be driven by richness but by large amounts of biomass, or dominance of one group. These relationships require more investigation, and may need to be incorporated into invasion theories.
Acknowledgements
Thanks to Bob Godfree for his encouragement, advice, and enthusiasm throughout the project, and for showing me just how interesting and fun the study of ecology can be. Much appreciation is extended to Brendan Lepschi and Dave Mallinson for their efforts in identifying many of the species, particularly grasses. Also thanks to Matt Woods for his help in the field, particularly amongst the heat and the flies! Finally I thank CSIRO and the CPBR for providing me with the chance to experience this valuable opportunity.
References
Elton, C.S. (1958) The ecology of invasions by animals and plants. Methuan & Co. Ltd, London.
Foster, B.L., Smith, V.H., Dickson, T.L. & Hildbrand, T. (2002) Invasibility and compositional stability in a grassland community: relationships to diversity and extrinsic factors. OIKOS, 99: 300-307.
Godfree, R., Chu, P.W.G. & Woods, M.J. (in press) White Clover (Trifolium repens) and associated viruses in the subalpine region of southeastern Australia: implications for GMO risk assessment.
Levine, J.M. (2000) Species diversity and biological invasions: relating local processes to community pattern. Science, 288: 252-254.
Naeem, S., Knops, J.M.H., Tilman, D., Howe, K.M., Kennedy, T. & Gale, S. (2002) Plant diversity increased resistance to invasion in the absence of covarying extrinsic factors. OIKOS, 91: 97-108.
Shea, K. & Chesson, P. (2002) Community ecology theory as a framework for biological invasions. Trends in Ecology and Evolution, 17: 170-176.
van Ruijven, J., De Deyn, G.B. & Berendse, F. (2003) Diversity reduces invasibility in experimental plant communities: the role of plant species. Ecology Letters, 6: 910-918
Wardle, D.A. (2001) Experimental demonstration that plant diversity reduces invisibility – evidence of a biological mechanism or a consequence of sampling effect? OIKOS, 95: 161-170.
Fern Distribution and Diversity in Australia
Samantha Citroen
Supervisor Dr Sophie Bickford
Introduction
Biodiversity analysis has become an increasingly important field as ‘hotspots’ of diversity are now becoming a targeted priority for conservation planning and preservation of natural areas. Data for all of the Australian fern flora has recently been made available through the electronic databasing of herbarium collection records of Pteridophytes. This gives an opportunity for the analysis of continental scale patterns of fern diversity. There are over 400 species of Australian ferns, of various ages and evolutionary lineages.
Species distribution patterns results from a combination of physiological tolerances, geological and evolutionary history, as well as interactions with climate. These factors were taken into consideration in the investigation of fern species distributions across the Australian continent. This project was an exploratory analysis into the fern distribution patterns and highlighted areas for further investigation. As well as providing a quantitative analysis of fern diversity, results from the project demonstrated the utility of herbarium data sets for large scale biodiversity analysis.
Aims
The main aim of this research was to determine continental distribution and diversity patterns of ferns in Australia, and analyse these patterns with respect to current environmental conditions and evolutionary history, to generate a hypothesis for fern distribution across the continent.
This project was the first of its kind in exploring spatial fern data from herbarium records. Thus included in its scope was distribution mapping of all Australian fern species and an assessment of the scale at which such data can accurately represent actual distribution patterns. This would provide a basis to assess its potential for range determination using other methods, such as bioclimatic envelope modeling.
Methods and results
Constructing the spatial database
Herbarium specimen records from eight of the Australian herbaria were collated and validated. These herbaria were Darwin, Brisbane, QRS, New South Wales, Canberra, Melbourne, Hobart and Perth and in total approximately 50 000 fern and fern allied group records were collated and databased. These records were then edited and validated to give a satisfactory dataset for analysis. This included editing the dataset to identify records that were duplicated, of poor geographic accuracy and taxonomic misnomers (synonyms, spelling errors etc.). This data was then entered into a GIS (ArcMap) and further edited for geolocational errors based on known species distributions and tolerances as described by fern experts*. The final edited dataset totaled around 40 000 records of 455 different fern species. Figure 1 shows all the records of fern collections that were used for the dataset. (*David Jones, Australia National Herbarium).
Spatial analysis of species richness, areas of endemism and biotic regions (areas with shared groups of species) were undertaken on this data by using a 50 X 50 km grid across the continental range and each cell (2500 km�) was analysed based on the species components.
Species richness and endemism
Species richness was calculated as the total number of different species in each cell. Counts ranged from 0 to 211 as a maximum count in the wet tropic region of Queensland (Figure 2). Areas of high species richness were identified as: Cape York (90*), Wet tropics (211*), Border Ranges (130*), Sydney (90*), Eastern Victoria (70*), Tasmania (65*) (Figure 2). (*maximum cell species richness count in cells of that area). Although we did not quantitatively analyse climatic relationships with species richness, there appeared to be a strong positive correlation with moisture availability and species richness.
Endemism was used to investigate the range restriction of species and to identify centres of endemism, and the data was analysed at different spatial scales for this. Endemism was calculated as Correct Weighted Endemism, the inverse of the range of a species summed for a cell over the total number of species in a cell. This was calculated at increasing spatial resolutions, using a increasing radius cell neighbourhood approach, so that localised areas endemism could be identified and compared to regional patterns. Cape York and the wet tropics appeared as areas of high endemism at the smallest scale. The south west of Tasmania was observed as an area of high endemism at a smaller scale, whereas all of Tasmania came up as an endemic area on a larger scale analysis. Figure 3 shows areas of endemism at the 3 cell radius scale of analysis.
Biotic regions
Areas with similar groups of species were determined using similarity analysis and a hierarchical classification process based on the species complements of each grid cell. Although further exploration of the species composition of these regions is required to fully analyse correlation patterns in distributions, from a preliminary analysis there appeared a strong correlation between fern distribution patterns and climatic zone distributions based on rainfall, humidity and temperature. Centres of endemism also appeared correlated with the different climatic regions. There was also an apparent connection with the southern group of fern flora and New Zealand species. Figure 4 presents the regions of similar species complements across the continent.
Evolutionary exploration
It was apparent that climatic gradients would not explain the totality of fern distribution across the continent and exploration into the question of whether there are remnant distribution patterns reflecting evolutionary histories.
The ferns were divided into two evolutionary groups; the Eusporangiate and Leptosporangiate groups. Species richness and endemism performed on distinctive groups. Correlation analysis indicated that there was no difference between distribution patterns of richness or endemism in the two groups, and this may be a result of the division having occurred very early in their evolutionary history. The ancient history of ferns have led to speculation that remnant Gondwanan patterns may still be apparent in their distribution. Despite searching the biogeographic, phylogenetic and palaeoecological literature we were unable to identify ‘Gondwanic’ elements at a relevant classificatory level from more modern elements, prohibiting separate spatial analysis of ‘Gondwanic’ elements.
Further exploration: Species range modelling
A preliminary investigation of the potential of BIOCLIM (ANUclim package) to predict fern species distribution from the dataset was undertaken. The results indicate that there is a potential for this method to predict fern distributions, although modifications to data and prediction accuracy are needed. (see figure 5)
Discussion and conclusions
This project successfully produced the first quantitative analysis of large scale patterns of fern distribution in Australia. As yet this analysis is considered preliminary, however it identified both patterns of distribution, in terms of species richness, endemism and fern species regions, as well as areas for further investigation. The data is still in need of further editing and analysis in terms of accuracy of records and analysis for distribution and collection density with regard to biodiversity. However the dataset was concluded to be appropriate for a continental scale approach to biodiversity analysis, but for smaller scale analysis a more comprehensive assessment of record accuracy would be required.
From preliminary analysis there appears a strong association between fern distribution and climatic gradients across the continent. Further investigation would give greater insight into the species patterns and centres of endemism and the factors contributing aside from environmental influences.
Acknowledgments
I would like to sincerely thank my supervisor, Dr Sophie Bickford (CPBR) for providing such a great experience, David Jones (CPBR) and Jim Croft (ANBG) for their assistance, advice and inspiration, Kim for his help in creating such a huge database, Shaun Laffan (University of Sydney) for his assistance in analysis and finally CPBR for providing the opportunity for me to participate in CSIRO research.
References
Barrington, D.S., (1993), Ecological and historical factors in fern biogeography, J. Biogeography, 20: 275-280
Bureau of Flora and Fauna (1998), Canberra, Flora Of Australia, Vol. 8. Ferns, Gymnosperms and Allied Groups, Australian Govt. Pub. Service.
Crisp, M.D., Laffan, S., Linder, H.P, and Monro, A. (2001), Endemism in the Australian Flora, Journal of Biogeography, 28: 183-198.
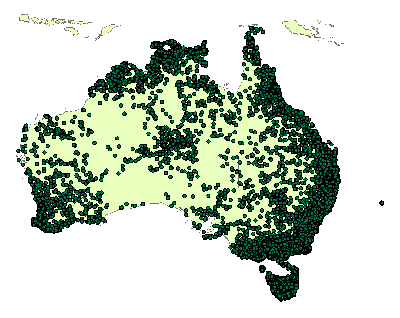
Figure 1 Fern species distribution records from herbarium records
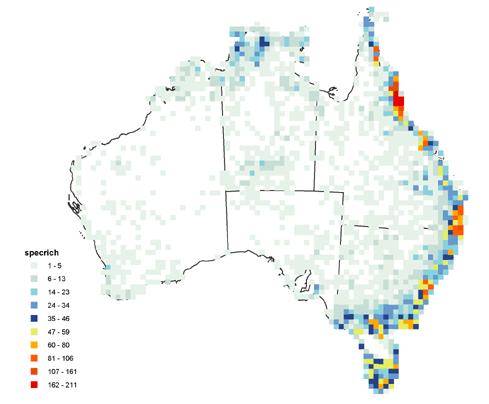
Figure 2 Ferns species richness in Australia
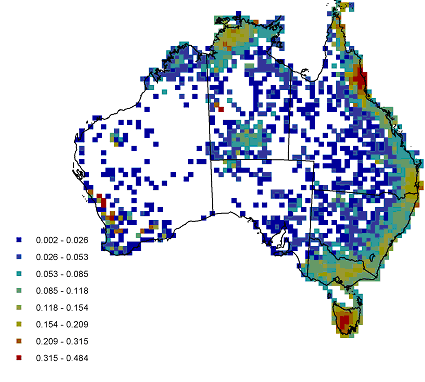
Figure 3 Areas of endemism of Australian fern species. Analysis conducted at the 3 cell radius scale.
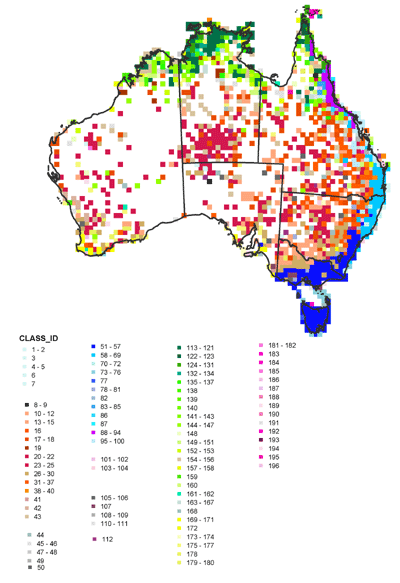
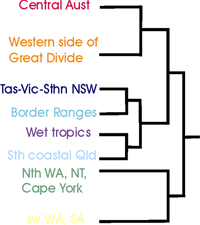
Figure 4 Biotic regions of fern species in Australia.
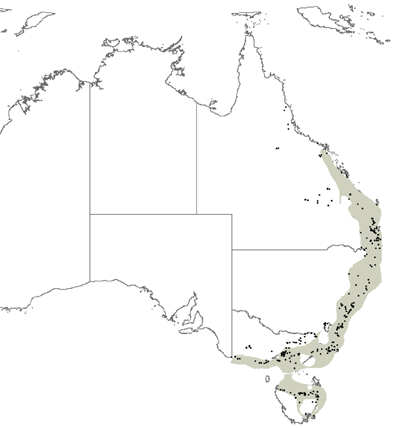
Figure 5 Prediction of the potential distribution of Cyathea dubia based on the bioclimatic distribution of herbarium specimen records.
********************
4. General Centre Matters
CSIRO Personal Assistants/Executive Assistants Conference – Report by Val Oliver
CSIRO Chief Executive Officer, Geoff Garrett, opened the two day conference with approximately 110 personal/executive assistants from CSIRO Divisions and Centres Australia-wide in attendance. The conference was held at Rydges Lakeside Canberra on 21-23 March 2004 and facilitated by Giuseppe Gigliotti from People Development, Corporate CSIRO and was
All participants attended a PA Empowerment Seminar where groups discussed issues they considered to be important for a PA/EA to carry out tasks efficiently.
Two workshops followed with group discussion on (i) writing a good rewards/double increment/reclassification case and (ii) making your objectives work at APA time. I chose the latter session where skills on streamlining objectives, setting new objectives and being proactive when considering tasks for the following year were improved.
Two PAs from the Division of Land and Water presented a seminar on administrative roles within the Division and the structure they work in. Participants agreed it was a unique presentation by the PAs and gave an excellent overview of the complexities of a large Division.
A variety of workshops were offered for the afternoon session, including Microsoft Office, Handling Outlook and Finance – PSS and Credit Cards. I attended the Handling Outlook session where participants contributed tips and hints to improve handling of MS Outlook. Tailored training courses are being proposed for MS Outlook and the information will be forwarded to PAs shortly for wider distribution to interested staff. Staff from CSIRO Information Technology were on hand to answer questions and provide smarter ways of using the program.
Warren King, Chris Yeats and Vivian Cateaux from CSIRO IT Manufacturing and Services provided an insight on working in a small team; what works for them in relation to advertising for staff, working productively when key staff are overseas and how office dynamics can play a critical part in communication.
A trade show by the conference sponsors provided up to date accommodation, conference and travel details for CSIRO staff. Qantas provided details on the New Domestic Fare Structure and New Fare Types and Features. This information is available from the Centre Office. A sample of sponsors included Avery, Hertz, Canberra Convention Bureau and most major accommodation providers.
Day 2 offered a selection of management and leadership skills workshops. I chose Coping with Change which considered the TEAM approach, i.e. Together Everyone Achieves More. The group discussed the importance of responding to what is happening in our environment and that communication is essential to change, i.e. one on one – not by email or phone. Coping with change differs between individuals and some key measures can assist, i.e. support/encouragement from manager, mutual respect, being accepted, being trusted, seek win-win situations, being included.
A scheduled seminar providing information on CSIRO Branding and Online Stationery by Corporate Communication staff was changed due to the illness of the presenter and, instead, a brief presentation gave an overview of the new electronic ordering system for CSIRO business cards and the plan to streamline the cards in line with One-CSIRO. Discussion sparked debate on the various methods being used across Divisions, which procedure was correct, the varied costs quoted by preferred printers and the procedure to be followed double sided card is required and dual logo requirements. Corporate agreed that some further work is required before a standard look and feel can be given to the cards and the feedback from the conference participants was valued.
Participants enjoyed lunch at the Division of Entomology’s BBQ area before the group departed on a choice of eight site and science tours including the Insect Collection, Wildlife Collection, Genetic Engineering, Australian National Herbarium, Homestead – Sustainable Ecosystems, Plant Biology, Weed Control and the Wind Tunnel. I participated in the tour of the Wind Tunnel facility at Land and Water’s Pye Laboratory where Bruce Stewart demonstrated the effects of wind on crops, smoke, forests and planes and the use of laser beams to measure smoke density/speed.
The conference concluded with afternoon tea and a tour of the Discovery Centre.
[Val Oliver]
********************
Executive Committee
The next meeting of the Executive Committee is scheduled for 28 April 2004.
[Val Oliver]