![An Australian Government Initiative [logo]](/images/austgovt_brown_90px.gif)


Truffle-like fungi in Australia
Terminology - truffle, truffle-like, false-truffle, hypogeous, sequestrate
What do they look like?
History of study of Australian truffle-like fungi
How many species are in Australia - and where do you find them?
Mycogeography of truffle-like fungi
Relationships
Evolution
Plants, animals and spore dispersal
Introduction
The truffle-like fungi differ from the bulk of the fungi discussed on this
website, in that the truffle-like fruiting bodies are generally produced underground
and have lost the ability to forcibly discharge their spores. The truffle-like
fungi have been better studied than many other fungal groups in Australia -
but 'better studied' does not mean 'thoroughly known'. ![]() Each
year, field work reveals more species and is exposing a surprising richness
of those fungi in this country.
Each
year, field work reveals more species and is exposing a surprising richness
of those fungi in this country.
At this point it's worth reminding you that macrofungal ascomycetes and basidiomycetes (SEE TWO MAJOR GROUPS SECTION) are the focus of this website, so only the ascomycete and basidiomycete truffle-like fungi will feature here. There are various other truffle-like fungi that won't be dealt with here. For those wishing to follow them further here are some genera which you can look for in the fungal literature or use in internet search engines: Acaulospora, Endogone, Gigaspora, Glomus. Collectively, these are part of a class of fungi called the Zygomycetes.
Terminology - truffle, truffle-like, false-truffle, hypogeous, sequestrate
Many terms have been used for the truffle-like fungi and you may come across some of these. Some people use the simple word "truffle" instead, but others hold this should be used only for the several species of European gourmet fungi that are sold as "truffles". Some would use the word "truffle" more broadly for any ascomycete species that was vaguely similar to the gourmet truffles and "false-truffle" for any basidiomycete equivalent. You will also see the word "hypogeous" - but this literally means "below ground", so it would be inappropriate for anything like Setchelliogaster, shown below, with its above-ground fruiting body. The term "sequestrate" is commonly used in the specialist literature and, unlike "hypogeous", is free of any connotations of the position of the fruiting body. However, on this website the expression "truffle-like" will be used as an all-encompassing term, simply because many people are familiar with the word "truffle", so using "truffle-like" immediately relates these fungi to a familiar concept.
What do truffle-like fungi look like?
The bulk of the truffle-like fruiting bodies are more-or-less spherical, though there is some variation in shape. They display a great variety of colours (for example - white, pink, yellow, orange, red, brown, purple), sizes (from a few millimetres to several centimetres in diameter) and consistency (from very soft to almost rock-hard).
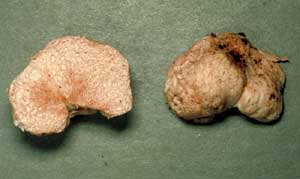 a 'squashed' globose Hydnangium carneum |
 a convoluted Peziza whitei |
Two more examples are a shortly stalked species of Setchelliogaster
sp. ![]() and a globose species of Tuber sp.
and a globose species of Tuber sp. ![]()
 |
 For some brighter colours here are Thaxterogaster porphyreus (left) and Paurocotylis pila (above) from New Zealand. |
As noted above, many of the truffle-like fruiting bodies mature underground and either never show above the surface or, at most, protrude just a fraction. Hydnangium and Tuber are examples of this. On the other hand, the Peziza, Setchelliogaster and Thaxterogaster are examples of truffle-like fungi with emergent or above-ground fruiting bodies, though as they are often less then a couple of centimetres in height or diameter, they are easily hidden by even a thin layer of leaf litter.
Why are these varied fungi bundled together and labelled "truffle-like fungi"? Apart from the position of the fruiting body in the soil, what is it that they have in common to justify such a grouping? The spores are produced internally, the spore-bearing tissue generally enclosed within an outer skin (called a peridium) and the spores are not forcibly released and rarely are they spread by wind or water - as is the case with the puffballs and their relatives. However, while they share various common features, the truffle-like fungi do not share a common ancestry. The evolution and relationships of the truffle-like fungi are discussed a little later.
There are both ascomycete and basidiomycete genera, but the outward form does not indicate which you have. Of the six examples pictured above - Hydnangium, Setchelliogaster and Thaxterogaster are basidiomycetes while Paurocotylis, Peziza and Tuber are ascomycetes. Ascomycetes and basidiomycetes are explained in the TWO MAJOR GROUPS section.
In the picture of Setchelliogaster you will see many small, empty chambers in the upper, expanded part of the fruiting body. The basidia line the walls of those chambers. In the ascomycetes Peziza whitei and Paurocotylis pila, the asci also line the walls of internal chambers, and this is the case in the bulk of the ascomycete species. Tuber is one of the exceptions, the asci being embedded in firm tissue.
History of study of Australian truffle-like fungi
As noted in the HISTORY SECTION, the 19th century saw Australian fungal collections sent to European mycologists for examination and publication and, given Australia's colonial history, it's not surprising that the bulk of that work was done by English mycologists. During the century the Europeans recorded a small number of Australian truffle-like fungi. The first monograph on Australian fungi was the 1892 Handbook of Australian Fungi, written by the English mycologist MC Cooke who commented that Australia had few subterranean species.
During the 20th century the number of species of Australian truffle-like fungi grew and grew and Cooke's statement was shown to be wrong. The reason for this explosion in knowledge was that there were now resident mycologists who were able to collect over extended periods (taking immediate advantage of good conditions) and examine their collections critically. The HISTORY SECTION gives brief details of the major resident mycologists.
Though the number of species grew steadily, for over half of the 20th
century both resident and overseas mycologists used superficial resemblances
to place Australian specimens into northern hemisphere genera. The move from
superficial resemblances to detailed microscopic, chemical and DNA studies has
changed fungal classification over the last 100 years and the classification
of the truffle-like fungi has changed dramatically since the 1970s. ![]() In
particular it became clear that, in many cases, the use of the names of northern
hemisphere genera for Australian specimens was inappropriate.
In
particular it became clear that, in many cases, the use of the names of northern
hemisphere genera for Australian specimens was inappropriate.
How many species are in Australia - and where do you find them?
About 300 truffle-like species are documented for Australia, but recent fieldwork has produced numerous collections of new species which are still awaiting publication. This work has also led to estimates of 1200-2400 as the eventual number of truffle-like species in Australia. By comparison, there are only a couple of hundred truffle-like species in Europe - an area that has been far more intensively studied than Australia.
They are found from rainforest to desert - with a couple of desert species eaten by Aborigines (see ABORIGINAL SECTION). As with Australian fungal exploration in general, fairly limited areas have been examined for the truffle-like species. The bulk of the collections have come from the south-west of WA, Adelaide and the surrounding hills, Tasmania and many parts of the Great Dividing Range along the east coast. In addition, the two desert truffle-like species have been collected from various scattered locations in central Australia and the north-west of WA. This still excludes most of Australia - areas such as the forests and woodlands of eastern Australia inland from the Divide, the drier areas of southern Australia and the bulk of the tropics.
While there certainly are rarely seen truffle-like fungi, as a group they're surprisingly common - even in suburban gardens. For example, during the transformation of the greater part of one suburban Canberra front yard (from a lawn and a few eucalypts to shrub-filled gardens beds), the digging up of the lawn areas revealed over a dozen species of truffle-like fungi (from about half a dozen genera) - including the Hydnangium and Setchelliogaster pictured above.
Mycogeography of truffle-like fungi
The study of fungal distributions is called mycogeography and a more detailed discussion of this topic is in the MYCOGEOGRAPHY SECTION. In this section there'll be a brief discussion of the relationships between Australian and non-Australian truffle-like fungi.
In Australia there are 83 genera of truffle-like fungi, though many of these are fairly recent discoveries (from the fieldwork referred to above) and not yet published in the scientific literature. Of those 83 genera, 42 are endemic. This is a remarkably high level of endemism at the generic level. By comparison, in the better studied areas of the northern hemisphere, North America has only 20 endemic genera (from a total of 77) and of Europe's 50 genera only 2 are endemic. Looking at other parts of the world, in New Zealand (with 30 genera of truffle-like fungi) there are 10 endemic genera, Africa has 8 endemic genera (from a total of 18), Asia has 8 from a total of 29 and South America has 9 from 23. As more of the southern hemisphere is explored, it is likely that the Australian level of generic endemism will decrease. For example, it is quite possible that some of the currently endemic Australian genera are of Gondwanan origin and therefore may be found in South America.
As noted above, about 50% of the truffle-like genera found in Australia are endemic but when it comes to species, the level of endemism is about 95%. Obviously there are many genera found in Australia, which are also found elsewhere - but for any such genus, the Australian species within the genus are almost always different to the overseas species in the same genus. The paper by Neale Bougher and Teresa Lebel noted that there were 51 genera found only in the southern hemisphere, 56 found only in the northern hemisphere and 49 that occur in both hemispheres. Of those last 49, many have fewer northern hemisphere species than southern hemisphere species.
Some of the southern hemisphere genera are widespread, showing a broad Gondwanan distribution and so being known from two or more of Australasia, South America, India and Africa. However, at the species level the distributions are generally more restricted. Australia and New Zealand share about 20 species - though six of these have been introduced from the northern hemisphere. Additionally, there are a few species with a natural distribution from Australia to south east Asia and India. Otherwise, species found naturally in Australia are not known from further afield (except where Eucalyptus has been introduced overseas). This contrasts with the situation in various other groups of fungi. For example, there are several polypore and mushroom species with a broader Gondwanan distribution (such as Australia and South America or Australia and southern Africa) but there are no truffle-like species with similar, broad southern hemisphere distributions. In the MYCOGEOGRAPHY SECTION, where you will also find examples of non-truffle fungi which are found naturally in both Australia and widespread areas of the northern hemisphere. When it comes to the truffle-like fungi, apart from the few species with a natural distribution from Australia through to Asia, there are no known truffle-like species found naturally in both Australia and the northern hemisphere.
The high levels of endemism (at both the generic and species levels) shows Australia to be a major centre of evolution of truffle-like fungi. Clearly, many of those genera have evolved in Australia, along with various specifically Australian plant and animal groups, after Australia separated from other land masses. Even where a genus is widespread (for example, of Gondwanan origin) the genus has obviously evolved considerably and independently in Australia to yield numerous specifically Australian species. The Australian truffle-like fungi are mostly mycorrhizal and have critical interactions with various native marsupials, a topic that will be dealt with below. Given these interdependencies, it is not surprising that so many endemic species and genera of truffle-like fungi have co-evolved with the various native animal and plant species.
 Melanogaster ambiguus |
The increase of human migration and trade over the past few centuries has resulted
in the deliberate or accidental spread of numerous organisms well beyond their
homes. While there are now strict quarantine laws in Australia, for over a century
after European settlement numerous animals and potted plants were brought here
- and with the plants came various fungi, including a small number of truffle-like
species. The genus Rhizopogon ![]() is common in pine plantations (it was a deliberate introduction for forestry
purposes, as were the bolete Suillus luteus and the mushrooms Amanita
muscaria and Lactarius deliciosus). Another example is Melanogaster
ambiguus. This specimen was collected in a plantation of Cork Oaks (Quercus
suber) in Canberra.
is common in pine plantations (it was a deliberate introduction for forestry
purposes, as were the bolete Suillus luteus and the mushrooms Amanita
muscaria and Lactarius deliciosus). Another example is Melanogaster
ambiguus. This specimen was collected in a plantation of Cork Oaks (Quercus
suber) in Canberra.
Relationships
At one time the bulk of the basidiomycete truffle-like fungi were placed in the gasteromycetes, which includes the puffballs, earthstars and several other groups where spores are produced inside the fruiting body.
It is now clear that many genera of fungi that produce above-ground fruiting
bodies have closely related truffle-like genera. For example the mushroom genus
Lactarius has the truffle-like Zelleromyces as a close relative.
Some inkling of this relationship is shown in the following pair of fungi, Lactarius
![]() and a Zelleromyces
and a Zelleromyces ![]() .
Note that when damaged, both exude a milky fluid. While this is an easily observed
feature, it wouldn't prove a relationship since such bleeding can be found in
a couple of other mushroom genera. However, both Lactarius and Zelleromyces
have colourless spores with surface ornamentations of ridges, warts or spines
that stain blackish-blue in an iodine solution. Spore features are important
in classification.
.
Note that when damaged, both exude a milky fluid. While this is an easily observed
feature, it wouldn't prove a relationship since such bleeding can be found in
a couple of other mushroom genera. However, both Lactarius and Zelleromyces
have colourless spores with surface ornamentations of ridges, warts or spines
that stain blackish-blue in an iodine solution. Spore features are important
in classification.
Here's another mushroom-truffle pair.
 Laccaria sp. |
Hydnangium carneum |
![]() On
the left is a species of Laccaria, a mushroom genus where many of the
species are a pinkish-red colour throughout and on the right is Hydnangium
carneum, a pinkish to pinkish-red truffle-like species. In this case there
is no oozing of liquid but look at these spore drawings: again Laccaria
on the left (with spores from two different species in the genus) and a Hydnangium
carneum spore on the right. The scale bar represents a hundredth of a millimetre.
On
the left is a species of Laccaria, a mushroom genus where many of the
species are a pinkish-red colour throughout and on the right is Hydnangium
carneum, a pinkish to pinkish-red truffle-like species. In this case there
is no oozing of liquid but look at these spore drawings: again Laccaria
on the left (with spores from two different species in the genus) and a Hydnangium
carneum spore on the right. The scale bar represents a hundredth of a millimetre.
While DNA studies are still to be done for many truffle-like genera, such studies
have been done for Hydnangium and show a close relationship with Laccaria.
|
Thaxterogaster porphyreus |
Here is Thaxterogaster porphyreus again, a New Zealand species
of truffle-like fungus. It has rust-brown spores, much like the mushroom genus
Cortinarius, an example of which is Cortinarius archeri ![]() .
It has been known for some time that Thaxterogaster and Cortinarius
are closely related, but obviously they're different because the spore-bearing
tissue of Thaxterogaster is enclosed while in Cortinarius the
caps open out completely to expose the spore-bearing gills to the air. Or is
that really an obvious difference?
.
It has been known for some time that Thaxterogaster and Cortinarius
are closely related, but obviously they're different because the spore-bearing
tissue of Thaxterogaster is enclosed while in Cortinarius the
caps open out completely to expose the spore-bearing gills to the air. Or is
that really an obvious difference?
One structural feature of Cortinarius and related genera is the presence
of a cortina. A cortina is a cobwebby veil ![]() that joins the stem to the cap margin and so covers the gills. It is present
in the young specimens, but is very flimsy and torn apart as the cap expands,
so that in fully mature specimens there are often few traces left and the spores
are unimpeded in their passage from the gills to the open air below the cap.
Dermocybe is related to Cortinarius and there are some odd Cortinarius
and Dermocybe species, such as Dermocybe globuliformis,
shown here.
that joins the stem to the cap margin and so covers the gills. It is present
in the young specimens, but is very flimsy and torn apart as the cap expands,
so that in fully mature specimens there are often few traces left and the spores
are unimpeded in their passage from the gills to the open air below the cap.
Dermocybe is related to Cortinarius and there are some odd Cortinarius
and Dermocybe species, such as Dermocybe globuliformis,
shown here. ![]()
This species is endemic to Australia and these pictures show mature specimens.
You will immediately notice several points - it certainly is mushroom-like (with
stem, cap and gills), but the stem is very short and the cortina is both quite
dense and persistent. While the spores are forcibly shot from the gills, in
the same way as in any ordinary mushroom, the dense cortina greatly hinders
the passage of the spores beyond the confines of the cap. However, even if there
were no cortina, the shortness of the stem means that the mature fruiting bodies
almost always remain buried in leaf litter. Here is a photo ![]() (taken on Black Mountain in Canberra) showing the fungus in a dry sclerophyll
habitat. The overlying leaf litter has been scraped away to show the mushroom
caps, hugging the soil. So, in contrast to the great majority of Dermocybe
species, the spores of Dermocybe globuliformis are not shed into
the air for wind dispersal. If that persistent cortina were a more membranous
veil it would virtually transform this mushroom into a proper truffle-like fungus.
(taken on Black Mountain in Canberra) showing the fungus in a dry sclerophyll
habitat. The overlying leaf litter has been scraped away to show the mushroom
caps, hugging the soil. So, in contrast to the great majority of Dermocybe
species, the spores of Dermocybe globuliformis are not shed into
the air for wind dispersal. If that persistent cortina were a more membranous
veil it would virtually transform this mushroom into a proper truffle-like fungus.
So you might begin to wonder just how enclosed a Cortinarius or Derrmocybe
can get before becoming a truffle. In fact, DNA evidence published in 2001 supports
the idea that the distinction between Thaxterogaster and Cortinarius
is artificial so that there should really be just the one genus, Cortinarius,
with a variety of fruiting body forms from mushroom to truffle-like. Dermocybe
globuliformis is a relatively recent discovery, its description having been
published in 1986. It was originally found in Western Australia, but is now
known to be widespread. A few other Cortinarius species with similarly
persistent cortinas (similar to that of Dermocybe globuliformis) are
known from North America and Kashmir. ![]() With
Thaxterogaster subsumed into Cortinarius, that genus would then
show a range of forms, from mushroom to "half-truffle" (like the example
of Dermocybe globuliformis) to fully enclosed truffle.
With
Thaxterogaster subsumed into Cortinarius, that genus would then
show a range of forms, from mushroom to "half-truffle" (like the example
of Dermocybe globuliformis) to fully enclosed truffle.
|
Peziza whitei |
In the case of the ascomycetes, at one stage the bulk of the ascomycete truffle-like
fungi were classed by themselves and thought to be quite separate from the ascomycetes
which released their spores forcibly. However, here also, more detailed research
has shown close connections between the truffle-like species and the "ordinary"
cup-like ascomycetes. For example, an ascomycete genus with some variety in
fruiting body form is Peziza. The bulk of Peziza species are disk-like
or shallow cups, such as Peziza retispora ![]() shown here. However, here again is Peziza whitei, shown here, a very
convoluted truffle-like fungus in the same genus.
shown here. However, here again is Peziza whitei, shown here, a very
convoluted truffle-like fungus in the same genus.
The relationships of some truffle-like fungi are still unknown. The fruiting
bodies in the genus Mesophellia ![]() have a layer of powdery spores between the tough outer casing and the "chewy"
centre. Here's another picture,
have a layer of powdery spores between the tough outer casing and the "chewy"
centre. Here's another picture, ![]() this time with the powdery spores having been tipped out. You can see a number
of struts between the outer casing and the central core. Currently this genus
is still kept with the puffballs, but that's largely by default rather than
because of overwhelming evidence.
this time with the powdery spores having been tipped out. You can see a number
of struts between the outer casing and the central core. Currently this genus
is still kept with the puffballs, but that's largely by default rather than
because of overwhelming evidence.
Evolution
As noted in the previous section, the microscopic and DNA evidence shows that, there are many closely related pairs of truffle-like and non-truffle-like fungi. A natural question is: In such closely-related pairs, did the truffle-like forms evolve from the non-truffle-like forms, was it the other way round or did both evolve from a common ancestor? This question has been debated for many decades, morphological evidence being used to support the different viewpoints. While there is still much work to be done it is now clear that the answer is: All of the above.
The recent molecular evidence strongly supports the idea that in a number of the mushroom/truffle-like pairs, evolution was from mushroom to truffle-like fruiting body. Similarly, in the ascomycetes the direction of evolution appears to have been from non-truffle-like to truffle-like.
There are also some truffle-like fungi with stinkhorns as their closest above-ground
relatives. An example is the genus Hysterangium ![]() ,
shown here. The species in Hysterangium often have a somewhat gelatinous
or rubbery internal texture. In the case of stinkhorn/truffle-like pairs, the
molecular evidence indicates that the above-ground stinkhorns evolved from truffle-like
ancestors.
,
shown here. The species in Hysterangium often have a somewhat gelatinous
or rubbery internal texture. In the case of stinkhorn/truffle-like pairs, the
molecular evidence indicates that the above-ground stinkhorns evolved from truffle-like
ancestors.
The truffle-like genus Rhizopogon ![]() is very closely related to the bolete genus Suillus
is very closely related to the bolete genus Suillus ![]() .
In fact, Suillus appears to be much more closely related to Rhizopogon
than to any other bolete genus. In this case, the DNA evidence suggest that
both genera evolved from a common ancestor.
.
In fact, Suillus appears to be much more closely related to Rhizopogon
than to any other bolete genus. In this case, the DNA evidence suggest that
both genera evolved from a common ancestor.
It is worth noting that the mushrooms release their spores forcibly (SEE MUSHROOM SPORE RELEASE SECTION). One early argument in favour of the mushroom-to-truffle direction of evolution was that, if evolution had been the other way, it would have been necessary for that complex spore discharge mechanism (common to all mushrooms) to have evolved independently a number of times. That seemed very unlikely. On the other hand, the stinkhorns release their spores passively (relying on fly-attracting chemicals) so truffle-to-stinkhorn evolution doesn't demand the development of any complex spore release mechanism. Moreover, the truffle-like fungi produce a wide array of chemical attractants, so the evolution to above-ground stinkhorns (and fly-attracting compounds) may not have required great changes to the chemical factories within the truffle-like ancestors.
Even the truffle-like fungi that are closely related to mushrooms are mostly
very un-mushroom-like, so it's easy to get the idea that the evolution from
mushrooms to truffle-like fruiting bodies would have been a complex process.
But that need not be the case. Even fungi that normally produce mushrooms can
occasionally produce aberrant, somewhat truffle-like forms. ![]() For
example, the species Lentinus tigrinus will at times produce a natural
mutant where the cap is closed-in and truffle-like. This aberrant form is caused
by a change in one gene. Technically, a recessive allele is involved.
For
example, the species Lentinus tigrinus will at times produce a natural
mutant where the cap is closed-in and truffle-like. This aberrant form is caused
by a change in one gene. Technically, a recessive allele is involved.
Truffle-like fungi, plants, animals and spore dispersal
Most truffle-like fungi are mycorrhizal, so the mycelia form symbiotic associations with plant roots. Mycorrhizal associations are essential for good plant health and there's more about this topic in the MYCORRHIZAS SECTION. Australia is rich in mycorrhizal fungi, in general, and field evidence suggests that the Australian truffle-like fungi form mycorrhizae with major forest trees such as Eucalyptus, Allocasuarina, Leptospermum, Acacia and Nothofagus. Taken as a group, these plant genera are widespread and at least one of these genera can be found in almost any Australian plant community. It is likely that, as is the case with other types of fungi, the truffle-like fungi form mycorrhizae with numerous other Australian plant genera.
 Long-nosed Potoroo (Potorous tridactylus) |
The truffle-like fungi rely mainly on animals for spore dispersal. The mature
fruiting body typically produces chemical compounds to attract animals to the
fruiting bodies and after the fruiting bodies have been eaten, the spores are
dispersed in animal droppings. Experiments have shown that the spores of various
truffle-like species remain viable after passing through animal guts. At least
30 species of Australian mammals have been found to eat fungi and, given the
lack of comprehensive studies, there may well be more animal species involved.
Of course, some of the known 30 may only be incidental eaters of fungi, for
there is still much work to be done on the levels of fungal consumption and
the types of fungi that are eaten. However, we already know that the majority
of the fungi involved, are truffle-like fungi. Many truffle-like fungi have
distinctive spores which can be found in droppings and identified (at least
to genus). The eating of fungi is well-developed in the rat-kangaroos (a marsupial
group which includes the bettongs and potoroos). The Long-nosed Potoroo (Potorous
tridactylus) of western Victoria eats over 40 different species of fungi,
the majority of which are truffle-like fungi. The rare Long-footed Potoroo (Potorous
longipes) of eastern Victoria and southeast NSW eats at least 33 species
and the Tasmanian Bettong (Bettongia gaimardi ![]() )
eats at least 49 species.
)
eats at least 49 species.
For Potoroos and Bettongs in general, fungi (mostly truffle-like) constitute at least 30-40% of the animals' diets throughout the year - but there is considerable variation between the species. For example, truffle-like fruiting bodies constitute 80-90% of the diet of the Long-footed Potoroo, regardless of season.
In the three-way (tree-fungus-marsupial) association it is clear that there
is a benefit to all parties. ![]() Tree
and fungus each benefit from the other through their mycorrhizal association.
The animal uses the fungus for food, but then helps disperse the fungal spores.
Tree
and fungus each benefit from the other through their mycorrhizal association.
The animal uses the fungus for food, but then helps disperse the fungal spores.
The truffle-like Phaeangium lefebvrei is found in the dry areas of north
Africa and the Middle East. In Kuwait the fruiting bodies are sought out at
by least 11 species of migrating birds and Bedouin hunters are reported to use
the fruiting bodies to bait bird traps. New Zealand has always lacked the small
mammals, that are so widespread in Australia, but has a number of ground-dwelling
birds occupying similar ecological niches. While there is still much to be learnt
about the New Zealand truffle-like fungi, a number of species produce very strikingly
coloured fruiting bodies - such as Paurocotylis pila ![]() and Thaxterogaster porphyreus
and Thaxterogaster porphyreus ![]() - and when these appear in the leaf litter of the forest floor, they look much
like fallen berries. It is possible that the various ground-feeding birds are
attracted to these colourful "berries" and act as spore-dispersal agents, so
fulfilling the role played by the various small mammals in Australia. Do some
Australian birds actively seek out truffle-like fungi? There certainly are colourful
truffle-like fruiting bodies and a number of berry-eating birds, especially
in the tropical and sub-tropical forests. Birds like the Cassowary and Brush
Turkey immediately come to mind.
- and when these appear in the leaf litter of the forest floor, they look much
like fallen berries. It is possible that the various ground-feeding birds are
attracted to these colourful "berries" and act as spore-dispersal agents, so
fulfilling the role played by the various small mammals in Australia. Do some
Australian birds actively seek out truffle-like fungi? There certainly are colourful
truffle-like fruiting bodies and a number of berry-eating birds, especially
in the tropical and sub-tropical forests. Birds like the Cassowary and Brush
Turkey immediately come to mind. ![]() While
a number of Australian birds have been seen eating fungi, and an Eastern Yellow
Robin has been happy to take truffle-like fungi flicked to it, the extent of
bird involvement in the spread of the spores of truffle-like fungi is unknown.
While
a number of Australian birds have been seen eating fungi, and an Eastern Yellow
Robin has been happy to take truffle-like fungi flicked to it, the extent of
bird involvement in the spread of the spores of truffle-like fungi is unknown.
Interesting pictures - though not Australian
 I have copied a plate from Magyarorsz ágföldalatti gombái , written by László Hollós and published in Budapest in 1911. The book's title can be translated as Hungarian underground fungi and it deals with species found in central Europe, so the plate seems out of place in an account of Australian truffle-like fungi but it does reveal more of the variety to be found amongst the world's truffle-like basidiomycetes. Moreover, there are native Australian species that are macroscopically similar to many of the illustrated species and at least two of those species are found in Australia. One is Melanogaster ambiguus, which has been introduced to Australia and is found associated with non-native trees. It is the subject of figures 31 to 33 and the last of those (a cross-section) is reproduced to the right of this paragraph. You can compare it with the photo of a sectioned Australian specimen that appears earlier on this web page. Figure 39 shows Hydnangium carneum, which also has been mentioned above. This is a native Australian species though the first published description had been based on specimens collected amongst Australian plants in Berlin. Hydnangium carneum is now known from numerous parts of the world where eucalypts have been introduced and this Hungarian book of 1911 had noted it as occurring already in England, Finland, France, Germany, Italy, Sweden and California as well as Australia.
I have copied a plate from Magyarorsz ágföldalatti gombái , written by László Hollós and published in Budapest in 1911. The book's title can be translated as Hungarian underground fungi and it deals with species found in central Europe, so the plate seems out of place in an account of Australian truffle-like fungi but it does reveal more of the variety to be found amongst the world's truffle-like basidiomycetes. Moreover, there are native Australian species that are macroscopically similar to many of the illustrated species and at least two of those species are found in Australia. One is Melanogaster ambiguus, which has been introduced to Australia and is found associated with non-native trees. It is the subject of figures 31 to 33 and the last of those (a cross-section) is reproduced to the right of this paragraph. You can compare it with the photo of a sectioned Australian specimen that appears earlier on this web page. Figure 39 shows Hydnangium carneum, which also has been mentioned above. This is a native Australian species though the first published description had been based on specimens collected amongst Australian plants in Berlin. Hydnangium carneum is now known from numerous parts of the world where eucalypts have been introduced and this Hungarian book of 1911 had noted it as occurring already in England, Finland, France, Germany, Italy, Sweden and California as well as Australia.
In that linked PDF, after the page of illustrations, you will find a page that gives the Latin species names and explanations, in Hungarian, of what each figure shows. The sentence near the top of the text explains that the specimens are shown life size and are drawings or paintings (unless noted as 'phot'). In the original publication the coloured, background rectangle measures 25 x 18 centimetres, which will help you work out the actual size of each specimen. Figures 4 and 5 show fruiting bodies with attached mycelial bundles and figure 35 (showing a cross section of a dried specimen) is based on an illustration produced by another author, Hazslinsky. The glossary given below will help you make sense of the remaining explanations. The words left unexplained are names of localities.
Hungarian-English glossary
és (and), fiatal (young), friss (fresh), két (two), kívülről (external view), közepén átmetszve (cross section - note that in figure 39 only the second word appears, but with the same meaning), összenőtt (stuck together), példány (specimens), száraz (dried), vén (old)