Centre for Australian National Biodiversity
Research
|
Centre for Australian National Biodiversity
Research
|
Supervisor: Dr Saul Cunningham (CSIRO Entomology)
 Introduction
Introduction Numerous studies have explored the ecological significance of body size. Of these, most studies have been conducted in the Northern Hemisphere and have mostly examined vertebrate species (Kruger and McGavin 2000). Physiological and ecological processes, for instance feeding and dispersal, have been linked to body size in birds and mammals (Brandle 2000), however, the relationships between body size and ecological processes for insects remain unclear.
Whilst knowing little about the relationships correlated with body size in insect communities, a few studies present findings which allow us to make some predictions about body size. For instance small body size may increase the dispersal abilities of insect populations (Basset & Kitching 1991, Gaston & Lawton 1988). For other body size relationships empirical investigation is required to qualify a theory like Basset & Kitching (1991) propose, which states, the more complex the habitat structure (fractal dimension), the smaller the body size of insects living there. Are there more small bodied insects dominating insect assemblages in structurally diverse forest?
Insect assemblages are incredibly diverse and most ecological work on insects to date has focused on describing the community structure in the context of species diversity. What has not yet been attempted is a comparative study that explores how insect community structure changes with different habitat types. The project aimed to explore the relevance of insect body size with regard to beetle assemblages occupying two distinct habitats.
 The
body lengths of 398 different beetle species collected from four (4)
different sites in South West Western Australia were measured and this data
was used to investigate the body size distribution of the beetle fauna. The
beetles were measured from the foremost part of the head to the tip of the elytra
or abdomen following the protocol outlined by Morse and Stork (1988). For the
data analysis body size data was natural log transformed from millimetres as
is common practice for this kind of ecological study.
The
body lengths of 398 different beetle species collected from four (4)
different sites in South West Western Australia were measured and this data
was used to investigate the body size distribution of the beetle fauna. The
beetles were measured from the foremost part of the head to the tip of the elytra
or abdomen following the protocol outlined by Morse and Stork (1988). For the
data analysis body size data was natural log transformed from millimetres as
is common practice for this kind of ecological study.
In this study three (3) collecting techniques were used:
Beetles were collected from two vegetation types at each of the four sites. The first vegetation type was Remnant forest which is characterised as being structurally diverse, rich in plant species and the most common tree species was Eucalyptus marginata (Jarrah). The second vegetation type was Plantation forest which is structurally simple and is essentially a monoculture with Eucalyptus globulus (Tasmanian Blue Gum) as the only woody vegetation cover
Remnant and plantation data was pooled from the 4 geographically separated sites to create one large sample in which the body size distributions of the beetle assemblages from the two habitats could be compared.
Because this area of research is novel and because little work has been done, the method was to explore data extensively and graphically before any analysis proceeded. With some basic questions in mind the first step was to determine what patterns occurred in the data. Once a pattern was found then the next step was to find or design a statistical test to test the pattern and continue the exploration of the data.
Rank - body size distributions:
The rank body size distributions for beetle species collected using both insecticidal fogging and malaise traps were very even, with body size declining in a smooth curve. The largest beetle, Conoderus sp. 2, was 21.5mm long and the smallest beetle, Anisomeristes sp. 2, was 0.64mm.
The average body size of beetles in plantations is larger than those in remnants. Remnant vs Plantation (insecticidal fogging fraction) (Np = 79, Nr = 194, test statistic t = 3.48 on 271 d.f., P = < 0.001). Considering malaise fraction data, the difference is in the same direction as the fog, but is not statistically significant.
Number of species per body - size class Histograms for Species occurring in the Remnant fraction only or the Malaise fraction only: Body size data (length) was natural log transformed for these histograms
Of significance was the difference in mean body size of the whole super-family Chrysomeloidea (leaf beetles), collected from remnants and plantations. Due to the family being very speciose the sample sizes were large enough (Np = 25, Nr =22) for a T-test (Test statistic t = 0.11 on 173 d.f., Probability = 0.912)
For the remnant malaise fraction, species occupying fewer sites were smaller on average than species occupying many sites.
There is a large spike in the 7.4 – 9.5 mm bin in the total abundance plantation fog vs body-size class that looks disproportionate to the rest of the graph. Weighted means were calculated before and after Gonipterus scutellatus (the most abundant species) was removed. Mean (w/ G.s. = 4.8mm, N = 3990) vs Mean (w/o G.s. = 3.6 mm, N = 2775). A single super-abundant species influences the mean body size in fog plantations.
There is a similar spike in the 7.4 – 9.5 mm bin in the total abundance remnant fog vs body-size class. Again weighted means were calculated and the presence of Gonipterus scutellatus influences the mean body size. Mean (w/ G.s. =3.42mm, N = 2204) vs Mean (w/o G.s. = 2.74mm, N = 1815). Removing Gonipterus scutellatus returns the distribution to normal and peaky.
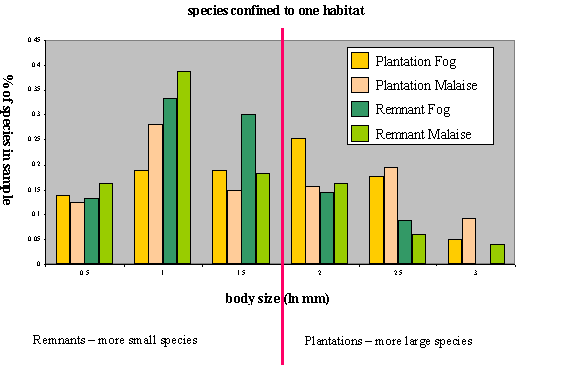
Figure 1. The number of species per body size class
distributions for beetles in remnant and plantation habitats. Each habitat has
beetles sampled from two different fractions, canopy knockdown with insecticide
or malaise trapping.
Basset, Y., Kitching, L., 1991: Species abundance and body length of arboreal arthropods associated with an Australian rainforest tree, Ecological Entomology, 16, 391-402
Brandle, M., Stadler, J., Brandl, R., 2000: Body size and host range in European Heteroptera, Ecography, 23, 139-147
Gaston, K.J., Lawton, J.H., 1988: Patterns in the distribution and abundance of insect populations, Nature, 331, 709-711
Kruger, O., McGavin, G.C., 2000: Macroecology of local insect communities, Acta Oecologica 21(1), 21-28.
Morse, D.R., Stork, N.E., Lawton, J.H., 1988: Species number, species abundance and body length relationships of arboreal beetles in Bornean lowland rain forest trees, Ecological Entomology, 13, 25-37.