![An Australian Government Initiative [logo]](/images/austgovt_brown_90px.gif)

|
|
 Birds
Nest and Cannonball Fungi
Birds
Nest and Cannonball Fungi These two groups of fungi have a superficial similarity. Both produce cup-like fruiting bodies that contain tiny "eggs", or peridioles. While each Cannonball fungus fruiting body contains just one peridiole, there are many in a Birds Nest fruiting body. However, despite the similarities in appearance the two groups are only distantly related and they release their spores in quite different ways. In this section you'll find out how the spore-containing peridioles are released.
There are several explanatory diagrams in this section. The particular colours that are used have no meaning and are used simply to help distinguish various features. The diagrams are also slight simplifications of reality, since their sole aim is to illustrate the important features.
|
|
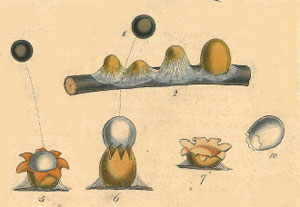
Cyathus novaezelandiae is an example of a Birds Nest Fungus. In the photograph you can see the egg-like peridioles in each fruiting body. The cups of the Birds Nest Fungi have a somewhat leathery consistency and each peridiole has a hard outer casing, within which there is a mass of spores. The cups vary in size but generally are between 5 and 10 millimetres in diameter.
In the case of Birds Nest Fungi, the peridioles are knocked out of the cups
by raindrops. When a peridiole lands somewhere - it sticks. In some genera of
Birds Nest Fungi the peridioles have sticky surfaces, but that's not the case
in the genera Crucibulum and Cyathus. In these genera the peridiole
has a long tail and the end of the tail is sticky and adheres to any surface
it comes into contact with. The photograph ![]() shows
some tailed peridioles from a specimen in the genus Cyathus. Each peridiole
is about 2 millimetres in diameter.
shows
some tailed peridioles from a specimen in the genus Cyathus. Each peridiole
is about 2 millimetres in diameter.
 Before
the peridiole is ejected, the tail is compressed into a small knob. The diagram
shows a peridiole (dark brown) and the knob (lighter brown), as well as an enlarged
view of the knob's contents. The knob has a thin casing, called the purse,
and this is the light brown area. Within the purse the tail (or funicular
cord) is compressed in a zig-zag fashion. The sticky end (or hapteron)
is shown in black and the rest in red. Furthermore, the lower end of the purse
is attached to the inner wall of the cup by hyphal strands (shown in grey).
Before
the peridiole is ejected, the tail is compressed into a small knob. The diagram
shows a peridiole (dark brown) and the knob (lighter brown), as well as an enlarged
view of the knob's contents. The knob has a thin casing, called the purse,
and this is the light brown area. Within the purse the tail (or funicular
cord) is compressed in a zig-zag fashion. The sticky end (or hapteron)
is shown in black and the rest in red. Furthermore, the lower end of the purse
is attached to the inner wall of the cup by hyphal strands (shown in grey).
When a raindrop forces a peridiole out, the grey hyphal strand initially stretches a little, but soon reaches its maximum possible extension and becomes taut. By now the peridiole has considerable momentum and the tautness of the hyphal strand results in the lower end of the purse being broken. This lets the peridiole continue its journey. There are two schools of thought as to what happens next.
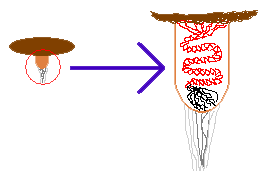
One school of thought holds that as soon as the purse is ruptured the cord
extends, so that the peridiole flies through the air trailing its tail. So
the entire sequence would be as shown in the accompanying diagram. In the
first figure you can see a blue raindrop falling towards the cup. In the second
figure the raindrop has hit the peridiole-filled cup and has dislodged one
brown peridiole. You can see it just beginning to leave the cup. Finally,
in the third figure, the peridiole has escaped the cup and is now trailing
a red tail with a black terminal segment.

The tail is made up of a number of hyphae and the black, terminal segment is sticky. As the peridiole flies through the air, the tail may swing about a bit and if the sticky segment hits something (such as a grass blade), it attaches itself straight away so that the flight of the peridiole stops. With the end of the tail now stuck, the rest of the tail just wraps around the grass blade and the peridiole comes to rest on that grass blade. Have you ever thrown a ball-on-a-string and then had it get caught in a tree? If so, you'd know just how well the string can wrap itself around a branch. Under hypothesis 1 a peridiole-on-a-tail would be very much like a miniature version of that ball-on-a-string.
The other school of thought holds that the tail doesn't immediately extend.
The peridiole flies through the air with just the sticky end of the tail exposed.
Now when the peridiole hits an obstacle the exposed sticky end could immediately
adhere to that obstacle. The peridiole would still keep moving since the examples
we are now looking at involve species with non-sticky peridioles. The continued
movement of the peridiole would now draw out the tail, which can extend considerably,
but after a very short time the peridiole would be reined in and brought to
a stop.

The splash-cup mechanism isn't restricted to Birds Nest Fungi. For example,
in the Plant Kingdom you can see similar splash-cups in liverworts of the genus
Marchantia ![]() . The circular splash-cups hold gemmae, which are small, spherical propagules
composed of a mass of liverwort cells. If a gemma lands in a suitable place
it will grow into a new Marchantia plant. The gemmae are not spores,
but are means of vegetative (or clonal) propagation. Each spherical gemma is
a bit like a miniature "cutting" from the parent. The Marchantia gemmae
cups are only a few millimetres in diameter. It is interesting to see that two
quite different (and unrelated) groups of organisms have, independently, come
up with the same solution to the problem of dispersing reproductive units.
. The circular splash-cups hold gemmae, which are small, spherical propagules
composed of a mass of liverwort cells. If a gemma lands in a suitable place
it will grow into a new Marchantia plant. The gemmae are not spores,
but are means of vegetative (or clonal) propagation. Each spherical gemma is
a bit like a miniature "cutting" from the parent. The Marchantia gemmae
cups are only a few millimetres in diameter. It is interesting to see that two
quite different (and unrelated) groups of organisms have, independently, come
up with the same solution to the problem of dispersing reproductive units.
While Birds Nest Fungi caught the eyes of various early botanists, it took a long time to discover the splash-cup method of peridiole dispersal. In 1686 the English naturalist John Ray thought that they were dispersed by rainwater which, once it had filled the cup, would overflow and so carry the peridioles with the overflow. In 1855 Julius Sachs cast doubt on the washing-out theory. Over three years he had observed some Birds Nest Fungi that grew on a small wooden bridge in the Canal Garden at Prague. This bridge was often completely under water during spring floods, but Sachs found no Birds Nest Fungi on another wooden bridge, about a hundred yards downstream. He had noted that the peridioles could float and that the spores germinated after being immersed. So, from the absence of the fungi from the other bridge, he concluded that the peridioles were not as easily washed out as had been supposed.
In between Ray and Sachs (and after) there were other hypotheses. Some supposed
that the peridioles were ejected by some elastic or spring mechanism and in
1877 one mycologist proposed that animals (presumably small ones!) sought out
peridioles for food and carried them out of the cups when the cords became entangled
in the animals' feet. ![]() The
first suggestion of the splashing mechanism came in 1927 when George W Martin
wrote that "...the peridioles could easily be spattered out of the cups by dropping
water into them from a pipette a couple of feet above them."
The
first suggestion of the splashing mechanism came in 1927 when George W Martin
wrote that "...the peridioles could easily be spattered out of the cups by dropping
water into them from a pipette a couple of feet above them."
However, the significance of Martin's observations didn't appear to sink in for some years. In 1941 AHR Buller commenced a series of dispersal experiments with Birds Nest Fungi in which he was soon helped by Harold Brodie, who continued the studies after Buller's death in 1944. These experiments showed that the raindrop mechanism is quite effective. The procedure was fairly simple:
Fix a Birds Nest Fungus fruiting body to a cork slab and put it in the middle of the laboratory floor.
Let water drops fall into the cups from a burette, positioned high up. The laboratory ceiling was high enough to allow the drops to fall 9 feet - thereby ensuring they were approaching terminal velocity by the time they hit the cup.
Search for peridioles on the laboratory floor and measure their distances from the cup. To make it easier to find the peridioles, spread white butcher's paper over the floor.
After experimenting with various species, the results were that the peridioles
splashed out for distances ranging from just a few centimetres to a little over
two metres. Mostly the distances were between 30 centimetres and a metre. You
can find out about these experiments (and much more about the Birds Nest Fungi)
in Brodie's book, given in the reference button. ![]() Brodie
also highly recommended standing out in a rainstorm, watching peridioles being
splashed out of Birds Nest Fungi! He also quotes an anonymous friend's thoughts
about the dispersal process: "Beautifully simple and simply beautiful.".
Brodie
also highly recommended standing out in a rainstorm, watching peridioles being
splashed out of Birds Nest Fungi! He also quotes an anonymous friend's thoughts
about the dispersal process: "Beautifully simple and simply beautiful.".
In 1843 JC Zenker described a new fungus on the leaves of Camellia plants.
Zenker didn't say where he'd found the plant and fungus, but presumably in Germany,
where he lived. This fungus produced small black pads, about two millimetres
in diameter, and Zenker gave it the name Leptostroma camelliae. Interestingly,
though Camellia plants were widely cultivated, there were no other reports
of this fungus turned up until 1933, in the United States. In that year some
specimens were sent to William Diehl of the US Department of Agriculture. Diehl
wrote that he was "...impressed by the startling reappearance of Zenker's species
after a lapse of nearly a century" and thoroughly investigated the new samples.
His conclusion: the black pads were nothing but the peridioles of a Birds Nest
Fungus. A re-examination of Zenker's published drawings showed evidence of the
cords on his pads as well. So, after 90 years, this mystery fungus was laid
to rest! ![]() Since
the peridioles can stick very firmly, they may often look like outgrowths, rather
than something stuck on. So Zenker's mistake is understandable.
Since
the peridioles can stick very firmly, they may often look like outgrowths, rather
than something stuck on. So Zenker's mistake is understandable.
|
|
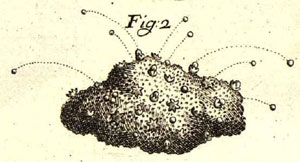
Whereas the Birds Nest Fungi are passive in their spore release and rely on raindrops to do all the work, the Cannonball Fungus (Sphaerobolus stellatus) is very active. When immature the whole fruiting body is a closed sphere. In the photograph you can see a line of immature fruiting bodies, going right (and slightly upward) from the centre of the photo.
 At
maturity the outer covering splits radially, from the apex of the sphere, leading
to the creation of a toothed cup, more-or-less hemispherical in shape. The number
of teeth vary between four and eight. If you look at the various open fruiting
bodies in the photo, you'll see that the cups do indeed have toothed (rather
than smooth) margins.
At
maturity the outer covering splits radially, from the apex of the sphere, leading
to the creation of a toothed cup, more-or-less hemispherical in shape. The number
of teeth vary between four and eight. If you look at the various open fruiting
bodies in the photo, you'll see that the cups do indeed have toothed (rather
than smooth) margins.
 Each
cup is about 2 millimetres in diameter and consists of a firm outer case and
an inner membrane - with a single reddish-brown peridiole sitting on that membrane.
The membrane is attached to the outer cup at the tips of the teeth mentioned
above. Within the cup, the peridiole sits in a bath of fluid. If you look at
the open cup near the lower left of the photo, you can see evidence of this
fluid in the way the flashlight has been reflected.
Each
cup is about 2 millimetres in diameter and consists of a firm outer case and
an inner membrane - with a single reddish-brown peridiole sitting on that membrane.
The membrane is attached to the outer cup at the tips of the teeth mentioned
above. Within the cup, the peridiole sits in a bath of fluid. If you look at
the open cup near the lower left of the photo, you can see evidence of this
fluid in the way the flashlight has been reflected.
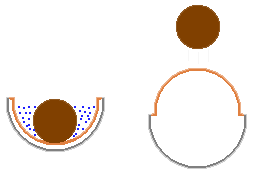
When the fruiting body is mature the inner membrane turns inside out and flicks
the peridiole into the air. The process is shown in the diagram, where there
is a grey outer cup, orange-brown inner membrane and dark brown peridiole. The
blue dots represent the fluid bath. The fluid in which the peridiole sits acts
as a lubricating fluid. It keeps the peridiole loose in the cup, so that it
is very easy to flick away.
Though the whole structure is only a couple of millimetres in diameter, the force of ejection is powerful enough to shoot the peridiole up to 6 metres away - so earning this fungus common names such as Cannonball Fungus or Artillery Fungus. Once the peridiole has been ejected, the inner membrane can collapse a little. In the photo you can see quite a few such collapsed membranes. The inversion of the membrane takes between a thousandth and a fifteen-hundredth of a second. The peridiole is shot off with an initial speed of around 3-5 metres per second. Unlike the peridioles in Birds Nest Fungi, the Sphaerobolus peridiole is fairly soft. It is composed largely of fatty materials. It compresses when it hits a solid object and stays firmly stuck.
 In
the first part of the Cannonball Fungus section, you've been told what happens
- the membrane turns inside out. You don't have to read any more if you are
quite satisfied with simply knowing what happens. The rest of this section goes
into a more detailed explanation of how it all happens and what roles the various
parts of the fruiting body play.
In
the first part of the Cannonball Fungus section, you've been told what happens
- the membrane turns inside out. You don't have to read any more if you are
quite satisfied with simply knowing what happens. The rest of this section goes
into a more detailed explanation of how it all happens and what roles the various
parts of the fruiting body play.
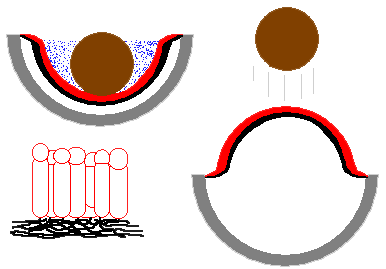
The accompanying diagram will help make sense of things. The figure on the
upper left depicts the firm outer cup (shown in grey), the inner membrane (shown
in red and black), an air layer (white) between the cup and membrane, the peridiole
(brown) and the fluid (blue stippling) in which the peridiole is immersed.
The inner membrane is composed of two layers. The red layer is formed by a palisade of mostly elongated cells, arranged radially (that is, oriented at right angles to the surface of the membrane), while the black layer consists of hyphae that are oriented more or less tangentially. The figure on the lower left shows a stylized, enlarged view of a small portion of the inner membrane, with the same red/black colour conventions.
The red layer of palisade cells absorbs water and so would expand laterally, but this expansion is held in check for a while and so builds up stresses. It is the release of these stresses that shoots the peridiole away.
One school of thought holds that though the red cells tend to expand, the non-swelling black hyphal layer holds the lower parts of the red layer firmly together. The other school of thought holds that the black hyphal layer plays no important role. Rather, the palisade cells are tightly packed and slightly pear shaped, broadest in the upper parts (which would swell more than would the lower areas).
Regardless of which explanation is correct, the result is the same. Stresses are created because within the inner membrane, the upper areas would expand more than the lower areas. While these stresses are building up, teeth in the outer cup continue to bend outwards, and so the outer cup continues to open. Eventually there comes a point when the outer cup has opened enough to allow the stresses to be released by the inner cup suddenly turning inside out - thereby flicking the peridiole out quite forcibly. This is shown in the figure on the right of the diagram.
Why do the palisade cells absorb water? At maturity there is a high sugar concentration within the cells and the cell walls are semi-permeable. The walls allow some substances (such as water) to move into or out of the cell - but numerous other molecules are unable to cross the barrier. Such semi-permeable cell walls are an important feature in all living organisms.
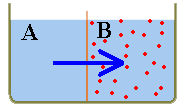 Suppose
you have two compartments (A and B), separated by a semi-permeable membrane.
Let both compartments be filled with water and add sugar (red dots) to B. It
is a fact of physics that water will be drawn from A into B. The same water
movement will occur even if some sugar is dissolved in A - as long as the sugar
concentration in A is less than the concentration in B. Such fluid flow (across
a semi-permeable membrane, from lower to higher sugar concentration) is called
osmosis. In the case of Sphaerobolus each of the palisade cells
is like compartment B, so water will be drawn in and the cells would expand
if possible. For the mechanism to work, a high level of humidity or damp surrounds
are necessary. Experiments show that a "loaded" Sphaerobolus won't fire
if it dries out.
Suppose
you have two compartments (A and B), separated by a semi-permeable membrane.
Let both compartments be filled with water and add sugar (red dots) to B. It
is a fact of physics that water will be drawn from A into B. The same water
movement will occur even if some sugar is dissolved in A - as long as the sugar
concentration in A is less than the concentration in B. Such fluid flow (across
a semi-permeable membrane, from lower to higher sugar concentration) is called
osmosis. In the case of Sphaerobolus each of the palisade cells
is like compartment B, so water will be drawn in and the cells would expand
if possible. For the mechanism to work, a high level of humidity or damp surrounds
are necessary. Experiments show that a "loaded" Sphaerobolus won't fire
if it dries out.
Before maturity, there are no sugars in the palisade cells. Instead, there is glycogen, a polymer composed of numerous simple sugar molecules joined together. However, glycogen has virtually no osmotic effect and so the swelling of the cells does not occur in the immature fruiting body. As the fruiting body opens and matures, the glycogen is converted into sugar molecules.
 Two
other important points are the air layer between the inner membrane and the
outer cup and that the membrane and cup are attached only at the teeth. Given
the air layer, there is no danger of the inner membrane sticking to the outer
cup. The fact that attachment is only at the teeth, means that the air layer
is in contact with the outside air. Consider the accompanying diagram, which
shows an eight-toothed Sphaerobolus fruiting body viewed from above.
You can see the peridiole on the red inner membrane, with the margin of the
outer cup shown in grey. Since the teeth are the only points of attachment between
the membrane and cup, in each shallow, V-shaped notch between neighbouring teeth
there is a gap (shown in white) between the inner membrane and the outer cup.
Two
other important points are the air layer between the inner membrane and the
outer cup and that the membrane and cup are attached only at the teeth. Given
the air layer, there is no danger of the inner membrane sticking to the outer
cup. The fact that attachment is only at the teeth, means that the air layer
is in contact with the outside air. Consider the accompanying diagram, which
shows an eight-toothed Sphaerobolus fruiting body viewed from above.
You can see the peridiole on the red inner membrane, with the margin of the
outer cup shown in grey. Since the teeth are the only points of attachment between
the membrane and cup, in each shallow, V-shaped notch between neighbouring teeth
there is a gap (shown in white) between the inner membrane and the outer cup.
These gaps allow the air layer between the membrane and cup to be in contact
with the outside air. When the membrane inverts, those gaps between the teeth
allow outside air to move quickly into the expanding space between the membrane
and cup. If the membrane were attached by all of its circumference, there would
be no gaps and the air layer between the membrane and cup would be cut off from
the outside air. In that case, the inverting membrane would be fighting against
a partial vacuum. ![]() That
would greatly slow the movement of the membrane and the peridiole would be shot
out only a short distance.
That
would greatly slow the movement of the membrane and the peridiole would be shot
out only a short distance.
|
|
The first description as well as the first illustration of this species appeared in Pier Antonio Micheli's Nova Plantarum Genera of 1729. He named it Carpobolus and the relevant plate is shown in this PDF (taken from the copy in the digital library of the Real Jardín Botánico, Madrid: http://bibdigital.rjb.csic.es/ing/index.php). Figures 1 and 2 show groups of fruiting bodies, some closed and some open with peridioles being ejected. The figures in the next two rows show entire, partial or sectioned fruiting bodies, with the two fruiting bodies marked A not as yet opened to expose the peridiole. The leftmost fruiting body has been partially cut to show the inner membrane labelled B. The letter E indicates the outer cup, F the inner membrane still curved downwards and K the same but now inverted. The final row shows peridioles and spores (called semina, or seeds, by Micheli). He noted that a peridiole or 'spherical fruit' (fructus sphaericus in his words) was packed with very fine seeds (ex seminibus minutissimus compactus in his words).
A detailed and beautiful coloured illustration appeared in Robert Kaye Greville's Scottish Cryptogamic Flora, a multi-part work that was produced during the 1820s in 72 fascicules. The Sphaeroblus plate was published in 1825 and is shown in this PDF. Figures 1 and 2 show groups of fruiting bodies, many still unopened. Figure 3 is a fruiting body shown in cross section just before it opens to expose the dark brown peridiole. Figure 4 gives an artist's impression of the peridiole in the process of being ejected and alongside it Figure 11 shows an inverted inner membrane after the peridiole has moved further away. Various views of whole or partial fruiting bodies are shown in Figures 5, 6, 7 and 10; peridioles in 8 and 9 (the latter with a small portion sliced off) and spores in 15. Figures 12 to 14 show cross-sections (12 - a fruiting body before dehiscence; 13 - a horizontal section, 14 - an enlarged portion of a horizontal section).
You can see scattered paint smudges in the particular copy of the plate shown in this scan. The illustrations in Greville's work were printed uncoloured and coloured in by hand, with 'master' coloured plates provided to the colourists as examples for them to follow. It is possible this was such a 'master' plate with the extra spots or streaks being colour trials by one of the colourists.