![An Australian Government Initiative [logo]](/images/austgovt_brown_90px.gif)

Spore discharge and dispersal in mushrooms
 Mushrooms
are basidiomycetes (SEE
TWO MAJOR GROUPS) with numerous basidia on each gill. A typical basidium
is a club-shaped structure, usually with four prongs at one end. Each such prong
is called a sterigma (with sterigmata the plural form) and the spores
develop at the tips of the sterigmata. Here is a stylized drawing of a basidium,
shown in green, with four brown spores. The colours in this diagram (and in
the ones to follow) have no significance and are simply used to help differentiate
the different structures. Moreover, the diagrams are stylized (rather than being
faithful depictions of actual specimens) but illustrate the important structural
features and principles involved.
Mushrooms
are basidiomycetes (SEE
TWO MAJOR GROUPS) with numerous basidia on each gill. A typical basidium
is a club-shaped structure, usually with four prongs at one end. Each such prong
is called a sterigma (with sterigmata the plural form) and the spores
develop at the tips of the sterigmata. Here is a stylized drawing of a basidium,
shown in green, with four brown spores. The colours in this diagram (and in
the ones to follow) have no significance and are simply used to help differentiate
the different structures. Moreover, the diagrams are stylized (rather than being
faithful depictions of actual specimens) but illustrate the important structural
features and principles involved.
The structure of a gill is illustrated and explained in detail in the (SEE TWO MAJOR GROUPS). Remember that most mushrooms have gills that are V-shaped in cross section (with the point of the V at the bottom of the gill) and are oriented vertically. The basidia are held out from the gill surface and protrude into the air space between two gills. Thus, throughout their development, the spores are exposed to the atmosphere between the gills. In order to explain the way in which the spores are ejected from the basidia it is necessary to look more closely at the spore and its attachment to the sterigma.
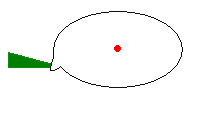 In
the diagram to the right the black, elliptical outline represents a mature spore
on the end of a sterigma (coloured green, on the left). Many spores are smooth
and ellipsoid, so this illustrates a fairly common situation - but the following
explanation holds for other spores as well. The red dot indicates the spore's
centre of mass. Note that the spore has a short, blunt, off-centre spike (called
an apiculus or hilar appendage) at one end and the spore is attached
to the sterigma at the apiculus. Every mushroom spore has an apiculus - though
there is some variation in size, shape and orientation of the apiculus between
species. The apiculus-sterigma boundary is a line of weakness and by the time
the spore is mature the link between apiculus and sterigma is very weak.
In
the diagram to the right the black, elliptical outline represents a mature spore
on the end of a sterigma (coloured green, on the left). Many spores are smooth
and ellipsoid, so this illustrates a fairly common situation - but the following
explanation holds for other spores as well. The red dot indicates the spore's
centre of mass. Note that the spore has a short, blunt, off-centre spike (called
an apiculus or hilar appendage) at one end and the spore is attached
to the sterigma at the apiculus. Every mushroom spore has an apiculus - though
there is some variation in size, shape and orientation of the apiculus between
species. The apiculus-sterigma boundary is a line of weakness and by the time
the spore is mature the link between apiculus and sterigma is very weak.
The discharge mechanism
Now it is a simple matter to explain the way in which the spore gets off the gill and away from the mushroom cap. The following diagrams illustrate the first part of the process (ejection from the sterigma) and the explanations follow.
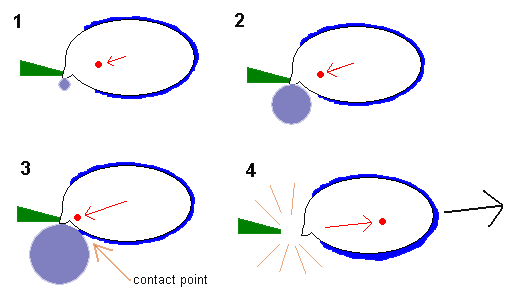
1. Between the gills the air is still and very humid. At the point of the apiculus the spore secretes a small amount of sugar molecules. If you've ever left an open sugar bowl on a kitchen bench for a lengthy time, you would have come back to find that the sugar had picked up some moisture from the air. Sugar is an excellent absorber of water vapour. Given the surrounding high humidity between the gills, water condenses onto the sugar at the tip of the apiculus and forms a drop (the small, solid, greyish-blue circle). This drop is known as Buller's drop. At the same time a thin film of water forms on a large area of the spore surface and this is represented by the brighter blue layer over much of the spore outline. The growing drop leads to a significant increase in mass at the apiculus, thereby causing the centre of mass to move towards the apiculus, as shown by the red arrow.
2 and 3. The water drop continues to grow in size as more water vapour condenses onto the drop's surface. This draws the centre of mass well away from its original point. Eventually the drop grows large enough to come into contact with the film of water on the spore surface. The contact point is arrowed. The drop may grow quite large in relation to spore size.
4. As soon as the drop comes into contact with the film, the drop collapses, with the water in the drop flowing into the watery film. This happens very quickly and the centre of mass moves very rapidly in more-or-less the reverse direction (as, again, shown by a red arrow). Simultaneously the spore is given considerable momentum, there is a break at the weak apiculus-sterigma boundary and the spore accelerates along the axis of the changing centre of mass (so moving off in the direction shown by the black arrow).
To give you some idea of the difference in speed, steps 1 to 3 are analogous to someone slowly stretching an elastic band and then, in step 4, the elastic is released so that it returns to its original size almost instantaneously. The momentum generated by the collapsing water drop is enough to give the spore an acceleration of 25,000 times the force of gravity. By comparison the NASA Space Shuttle has a maximum acceleration of just a few times the force of gravity. The spore loses about 1% of its mass in the secretion of the sugars on the apiculus. To continue the rocket comparison, the Space Shuttle uses about 50% of its own weight in fuel during the first two minutes after launch.
After discharge - getting the spores further away
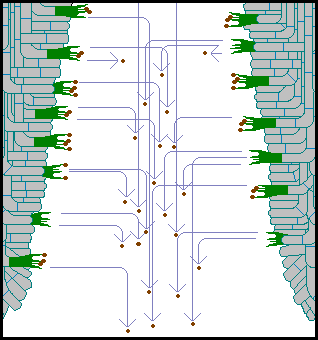 While
the spore leaves the basidium with a tremendous acceleration, it is small and
quickly feels the effects of air resistance. The spore briefly follows an almost
straight-line path away from the basidium, then slows, loses the forward momentum
given by the initial acceleration and finally drifts down (under the influence
of gravity) in the air gap between the gills until clear of the cap - where
even the slightest of air currents will carry the spores further afield. In
the following diagram the blue lines show the paths of a number of spores, some
just released from the basidium and others nearly beyond the bottom of the gills
and into the open air.
While
the spore leaves the basidium with a tremendous acceleration, it is small and
quickly feels the effects of air resistance. The spore briefly follows an almost
straight-line path away from the basidium, then slows, loses the forward momentum
given by the initial acceleration and finally drifts down (under the influence
of gravity) in the air gap between the gills until clear of the cap - where
even the slightest of air currents will carry the spores further afield. In
the following diagram the blue lines show the paths of a number of spores, some
just released from the basidium and others nearly beyond the bottom of the gills
and into the open air.
Once the spores have cleared the bottom of the cap, air currents carry them
away. But even at the bottom of the cap there is a danger to overcome. Should
a gently falling spore be exposed to the prevailing wind immediately after clearing
the bottom of the cap, there is a risk of it's being blown back onto the bottom
edge of a gill and so getting no further afield. Wind tunnel experiments have
shown that immediately beneath the cap there's a narrow band (about 2-3 mm deep)
where the wind speed is significantly lower than the incident wind speed. Below
that band is a zone of greater wind speed and near the ground there is a boundary
layer of calm air. On the leeward side of the cap there is always turbulent
airflow.
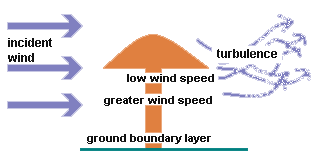 Thus
the spore doesn't feel the full effect of the surrounding wind speed immediately
after leaving the protection of the cap, so allowing more vertical movement
to the spore before being subject to a dramatic wind-induced, horizontal acceleration.
While the evidence suggests that this will prevent (or at least reduce) the
incidence of spore blow-back onto the gills, that conclusion is still to be
confirmed. Once the spores are a few millimetres away from the cap they can
be picked up by the faster winds and carried considerable distances.
Thus
the spore doesn't feel the full effect of the surrounding wind speed immediately
after leaving the protection of the cap, so allowing more vertical movement
to the spore before being subject to a dramatic wind-induced, horizontal acceleration.
While the evidence suggests that this will prevent (or at least reduce) the
incidence of spore blow-back onto the gills, that conclusion is still to be
confirmed. Once the spores are a few millimetres away from the cap they can
be picked up by the faster winds and carried considerable distances.
The wind-tunnel studies also showed that taller conical or bell-shaped caps showed the greatest reduction in wind speed below the cap. Interestingly, some common species of exposed windy, grasslands produce such caps.
Of course, changes in wind speed and direction (during the descent of the spores)
as well as interactions between the wind and nearby obstructions such as plants,
rocks and fallen twigs will obviously affect the spore paths. For example, you
will often see noticeable spore deposits on the ground beneath mushrooms - showing
spores which did not get far away. ![]() However,
while the wind-tunnel experiments will often reflect ideal (rather than natural)
settings, such experiments do show that there is more to mushroom architecture
than you might first suppose.
However,
while the wind-tunnel experiments will often reflect ideal (rather than natural)
settings, such experiments do show that there is more to mushroom architecture
than you might first suppose.
More about mushroom growth - and other ballistosporic basidiomycetes
 In
the bulk of mushroom species the spores in different parts of a gill may mature
at the same time. The spores near the bottom edge of a gill may mature at the
same time as those at the top of the gill. So, at any given time, many different
areas of a gill will be releasing spores into the surrounding air. This was
shown above, in the diagram of spore trajectories between two gills. The vertical
orientation of the gills is therefore critical, to maximise the number of spores
that get beyond the confines of the cap. For example, the diagram (right) shows
two, dramatically non-vertical grey gills. Any spore that begins the vertical
part of its trajectory in the area shaded brown will not get beyond the cap,
but will be trapped on the right hand gill. Spores are sticky, so once a spore
lands on the opposite gill, it won't get any further.
In
the bulk of mushroom species the spores in different parts of a gill may mature
at the same time. The spores near the bottom edge of a gill may mature at the
same time as those at the top of the gill. So, at any given time, many different
areas of a gill will be releasing spores into the surrounding air. This was
shown above, in the diagram of spore trajectories between two gills. The vertical
orientation of the gills is therefore critical, to maximise the number of spores
that get beyond the confines of the cap. For example, the diagram (right) shows
two, dramatically non-vertical grey gills. Any spore that begins the vertical
part of its trajectory in the area shaded brown will not get beyond the cap,
but will be trapped on the right hand gill. Spores are sticky, so once a spore
lands on the opposite gill, it won't get any further.
The possession of V-shaped gills also means that the air gap between neighbouring gills increases towards the bottom of the cap. While mushrooms do not sway greatly in the wind, they are not rigid structures. The increasing air gap gives the spores a better chance of escape, should the mushroom be tilted slightly (with the gills therefore no longer vertical).
During growth of the mushroom, the stem grows upwards, against gravity. It is necessary for the cap to be raised high enough above the still, surface boundary layer and any obstructions so that the falling spores can be dispersed by air currents. The gills also respond to gravity, but in the opposite way to the stem. Should something be not quite right with the cap orientation, the developing gills can make some corrections to ensure their proper orientation. In the bulk of mushroom species there are strong developmental controls aimed at ensuring that vertical gill orientation.
A spore that is shot off the basidium in the way described above is called a ballistospore. When a spore is shot off the basidium on the gill of a mushroom, it is important that the force isn't strong enough to send it to the neighbouring gill, for the spore would remain stuck there. On the other hand, the force must be sufficient to get the spore a reasonable distance away from the basidium, so that it doesn't get trapped on the gill it started from. While there is some variation in the distances that the spores of a specific mushroom species are ejected, the distances are all in a fairly narrow range. However, there is considerable variation in the ranges between species, with some species ejecting the spores no more than a tenth of a millimetre while others may shoot them out to half a millimetre.
Mushrooms are not the only basidiomycetes with ballistospores, for the same mechanism is found in various other types of basidiomycete fruiting bodies - boletes, the polypores, corticioid fungi, jelly fungi, coral fungi and stereoid fungi. Basidiomycetes such as puffballs, stinkhorns and the truffle-like species are "passive" spore releasers, without ballistospores.
The Inkcap mushrooms
Coprinus comatus, a common and widespread mushroom-producing fungus, is commonly called the Shaggy Inkcap, because the mushroom cap has prominent white to pale brownish scales that stick out from the cap (giving it a shaggy appearance) and the cap dissolves into an inky mess and drips away. The young caps of this mushroom resemble tall, rounded cones. The mushrooms are often up to 15 centimetres tall and found in lawns and along roadsides. This stylized diagram shows it in cross-section, with the gills in grey, the stem dark brown and the thin flesh of the cap lighter brown
 Coprinus comatus above and right |
 |
 |
The photo on the right shows Coprinus comatus with about half the cap already dissolved. Eventually, all that will remain, is the stem and a small, inky disk of flesh at the top of the stem. You can use that inky mess from a Coprinus comatus as ink. The picture below shows a blank herbarium label with some pre-printed headings in ordinary printers' black ink. The paler ink is from Coprinus comatus.

Question: Why does the cap dissolve and drip away? Answer: To help spore dispersal.
The densely packed gills of Coprinus comatus are parallel-sided, rather
than with a V-shaped cross-section, and without the strict vertical control
of gills that is found in the bulk of mushrooms. This would seem to make effective
spore dispersal very difficult. However, Coprinus comatus has overcome
these difficulties and the following diagram (in the same style as the earlier
diagrams) shows the process.
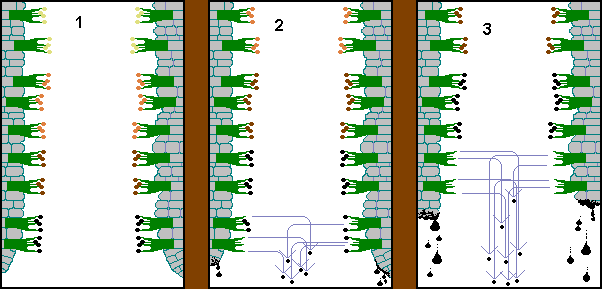 Figure
1 shows parts of two neighbouring Coprinus comatus gills, in stylized
cross-sections. Unlike the great majority of mushrooms the spores of Coprinus
comatus mature in a regimented way. The spores near the lower edge of the
gill mature first. The spores are colourless when immature but then pass through
various shades of pink and brown to become black at maturity. Figure 1
shows this, with the lowest basidia bearing black spores and a lightening of
spore colour as you look up the length of a gill - reflecting differing stages
towards maturity.
Figure
1 shows parts of two neighbouring Coprinus comatus gills, in stylized
cross-sections. Unlike the great majority of mushrooms the spores of Coprinus
comatus mature in a regimented way. The spores near the lower edge of the
gill mature first. The spores are colourless when immature but then pass through
various shades of pink and brown to become black at maturity. Figure 1
shows this, with the lowest basidia bearing black spores and a lightening of
spore colour as you look up the length of a gill - reflecting differing stages
towards maturity.
The mature spores that were shown in figure 1 are ejected by the water-drop mechanism and figure 2 shows the spore trajectories. At the same time the spores in the next band of basidia are now mature. You can see that the spores that were dark brown in figure 1 are now black - and there have been other colour changes in the spores higher up. Figure 2 also shows the lower edges of the gills beginning to dissolve into an inky mess which drips away.
In figure 3 more of each gill (including the basidia which shot off their spores in figure 2) has disappeared as ink drops and now the second band of basidia is releasing its spores.
This process continues, the spores maturing in rising bands and the gills dissolving and dripping away below the bands of maturing spores. Each group of mature spores therefore has only a very short distance to fall, before being clear of the ever-diminishing cap and so the lack of V-shaped gills is no hindrance to spore dispersal.
The absence of V-shaped, vertically oriented gills does not, in any way, make Coprinus comatus an "inefficient" mushroom. It is a prolific producer and disperser of spores, and has simply evolved a different (and effective) solution to the problem of spore dispersal.
The members of the genus Coprinus are commonly called Inkcaps, because the caps dissolve into an inky mess and drip away. The caps of many of the other Coprinus species do not dissolve quite as dramatically (and with such copious ink) as those of Coprinus comatus. Moreover, Coprinus comatus produces the largest mushrooms in the genus, with most Coprinus species producing mushrooms with caps no more than two to three centimetres tall.
What gives the spore its momentum? Some technicalities.
The bulk of what was said above was devoted to describing "what" happens during spore release. This section discusses some of the basic physics involved in the "surface tension catapult" mechanism that has already been described. It adds nothing new to "what" happens but briefly explains "how" it happens.
As the drop expands, its surface area increases and so does the total surface energy arising from surface tension. When the drop comes into contact with the film of water, the drop collapses, with the water from the drop flowing into the watery film. That action reduces the total surface area of the water - and so also the total surface energy. Conservation of energy is a fundamental law of physics. Energy never disappears, though it may be transformed from one form into another. So, if you start with a certain amount of energy you must end up with the same total amount - though it may appear in different forms.
During the spore launch, while the total surface energy is reduced, the spore (and its watery coating) gain kinetic energy (and momentum). Basic physics indicates that some of the original surface energy would be transformed into heat - though how much, is unknown.
The sterigma also plays an important role in ballistospore discharge. Throughout
the process the sterigma maintains its shape by internal turgor pressure. The
sterigma therefore provides a rigid launching platform for the spore and is
able to absorb the recoil from the accelerating spore, with only very minor
deformation. ![]() Without
that rigidity, energy would be wasted in deformation of a laxer sterigma - so
leaving less energy to contribute to spore acceleration. In an analogous way,
you can jump higher if you launch yourself from solid ground - rather than from
a much more compressible surface.
Without
that rigidity, energy would be wasted in deformation of a laxer sterigma - so
leaving less energy to contribute to spore acceleration. In an analogous way,
you can jump higher if you launch yourself from solid ground - rather than from
a much more compressible surface.