![An Australian Government Initiative [logo]](/images/austgovt_brown_90px.gif)

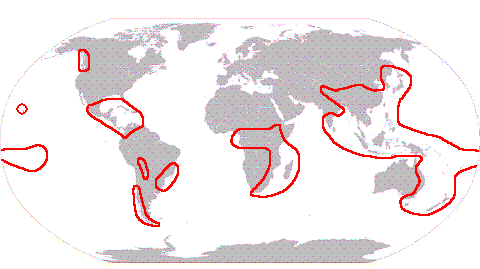
Case StudiesBryogeography of the HypopterygiaceaeThe moss family Hypopterygiaceae contains eight genera. This page discusses the bryogeography of the family with hypotheses as to how some the family's current distribution arose. The bulk of the information is taken from Chapter 7 of Kruijer's monograph listed in the references at the end of this page. The genus Hypopterygium, on which the family name is based, contains seven species and, as a genus, is widespread. The following map shows the parts of the world in which the genus is found.
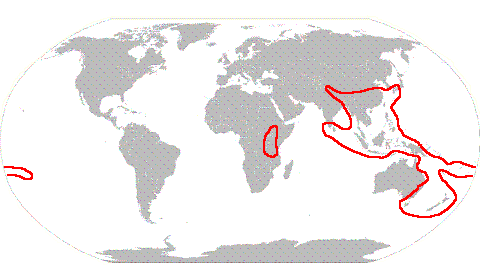
To say that the genus is found in all those areas is not to say that all species are widespread. Hypopterygium tamarisci is very widespread and found in most of the areas marked above, except Hawaii and the north-west of North America. By contrast, Hypopterygium sandwicense is known only from Hawaii. Cyathophorum is another genus with seven species and has the following distribution.
The combined distribution of the two species of Lopidium covers much of the area occupied by Cyathophorum but with a presence in more of continental Africa as well as Madagascar and parts of South America. Each of the remaining five genera contains only one species. Dendrocyathophorum is found from Papua New Guinea through to SE Asia, China and Japan. Arbusculohypopterygium is found only in South America while Canalohypopterygium, Catharomnium and Dendrohypopterygium are restricted to New Zealand and some adjacent islands. The spores of the Hypopterygiaceae generally have diameters in the 9-25 micrometre range, though sometimes up to 35 micrometres. In theory the bulk of the spores of the Hypopterygiaceae are small enough to be carried several thousand kilometres by air streams. The vegetative propagules are larger and capable of only short distance dispersal. Spores of several members of the Hypopterygiaceae were included in experiments on UV tolerance and resistance to frost and drought. The experiments indicated that some of those Hypopterygiaceae could survive transport between, say, Australia and New Zealand, during winter in streams of moist air. However, the spores would be very unlikely to survive a trip between Australasia and South America. Overall, long-distance dispersal would appear to be rare in the Hypopterygiaceae so that the diversity in, and distribution of, the family has been brought about by a combination of plate tectonics and continental drift. The family is of Gondwanan origin. There are several pieces of evidence that support this hypothesis. Research supports the idea that the genera Arbusculohypopterygium, Canalohypopterygium and Dendrohypopterygium are ancient members of the family and these are endemic to New Zealand or South America. There is a high diversity at both the generic and species level in New Zealand and at the species level in Australasia and Indo Malaysia. Much of the family's present distribution is on Gondwanan fragments. Regarding a more precise location for the family's origin, the number of New Zealand endemics suggests that the family originated in eastern Gondwana. Before Gwondana broke up New Zealand was part of eastern Gondwana and was located near what is now Marie Byrd Land in Antarctica. The evidence suggests that the ancestral Hypopterygiaceae were probably found in active mountain ranges on the continental margin, in cool, temperate conditions and occupied humid, terrestrial habitats at low altitudes. Today the terrestrial Arbusculohypopterygium, Canalohypopterygium and Dendrohypopterygium are largely found in such habitats, though occasionally at higher altitudes. Ancestors of the ancient genera now found in New Zealand must have been in what is now New Zealand before its separation from Gondwana. Since the New Zealand micro-continent separated from Gondwana between 84 and 86 million years ago the family Hypopterygiaceae would be at least 84 million years old. As the fragments of the original Gondwanan home of the Hypopterygiaceae drifted their separate ways species evolved independently in each fragment. Over time increasing areas of the Gondwanan fragments moved into warm-temperate zones and the descendents of the originally cool-temperate Hypopterygiaceae moved into higher altitudes. The move into warm-temperate zones, with the consequent higher rainfall and humidity meant that more exposed habitats could be occupied and descendents of the terrestrial species now colonized plants and rocks as well. The increase in angiosperm diversity also created a variety of woody substrates. The gametophytes of the three ancient genera are dendroid, with a fan of leafy branches atop a robust stem that is several to many centimetres tall. Such a growth habit is advantageous to terrestrial bryophytes growing on the forest floor, since the photosynthesizing leaves are kept above the terrestrial leaf litter layer. Such stems confer no advantage to bryophytes growing on boulders or tree trunks. Amongst the species that evolved later are non-dendroid forms, with gametophytes that are fan-like or just simple leafy stems. In his monograph Kruijer gives possible hypotheses for the present distribution of each species. It would take too much space to include all those hypotheses on this web page, so a few examples will suffice. The Hypopterygiaceae would have reached Asia once Gondwanan fragments had migrated close enough to make the dispersal distance no longer a challenge. There are two possibilities as to migration routes into Asia. One is that ancestors of the current Asian Hypopterygiaceae were carried north on the Indian subcontinent and then moved further into Asia once India collided with Eurasia. Another possibility is that those ancestors island-hopped north from Australasia. Included in the current distribution of Hypopterygium tamarisci are the Neotropics, southern South America and Africa. During the early Cenozoic period the northern part of South America was close to western Africa and this species may then have reached the Americas from Africa. Alternatively, the species may have reached southern South America from the east Gondwanan Hypopterygiaceae homeland via Antarctica and then migrated north. There is insufficient evidence to decide between the two hypotheses. Of the two Lopidium species, Lopidium concinnum occurs in Australia, New Zealand, Chile, Bolivia and Brazil and Lopidium struthiopteris is found in Australia, Africa, southern India, Sri Lanka, many areas of South-East Asia, southern China, Japan, Melanesia and Polynesia. In Australia the genus is found in Tasmania and, on the mainland, in eastern Australia between the coast and the Great Dividing Range. However, the only place where the Australian ranges of the two species overlap is near the border between New South Wales and Queensland. Otherwise, in Australia, Lopidium concinnum is found to the south and Lopidium struthiopteris to the north. In Kruijer's view the current world distributions of the two species reflect the occurrence of Lopidium concinnum in the cooler southern areas in Cretaceous Gondwana and the other's occurrence in the warmer north at that time. Hypopterygium sandwicense occurs only in the Hawaiian islands, more precisely on the islands of Hawaii and Maui and is reported to be rare on the latter. Presumably propagules of an ancestor reached the islands. Earlier in this page it was said that long distance dispersal was probably rare in the Hypopterygiaceae but rare does not mean impossible. Over a long period of time the chance of a rare event happening increases. References
|