
Spider Orchids
A genus of terrestrial orchids which grow in a range of forested and shrubby habitats. The plants are hairy on most parts. The sterile plants have a single hairy leaf, often purplish at the base. The flowering plants have a single hairy basal leaf and a hairy flower stalk, which is one to four flowered. The brightly variably coloured spidery flowers range in size, measuring 1-12 cm in width. The spreading to drooping sepals and petals are similar in size and shape, with numerous small glands on the tips. The labellum has elongate to triangular marginal teeth, and is adorned with hairs and calli, which are arranged mostly in two or more rows. The winged column has a pair of conspicuous yellow glands at the base. Arachnorchis flower in spring and early summer with one species flowering in winter. They occur in all states throughout Australia except the Northern Territory.
Similar Genera
Jonesiopsis
Significant Generic Characters
Arachnorchis is a terrestrial genus characterised by tubers partially enclosed in a fibrous sheath; reproduction from seed; single relatively broad hairy leaf usually purplish at the base; thin wiry hairy scape; leaf trichomes eglandular, multicelled with an enlarged long white basal cell; 1-4-flowered inflorescence; moderately small to large, often spidery, resupinate, green, white, pink, red or yellow flowers; sepals and petals subsimilar, attenuate, the tips extended into caudae or thickened into club-like units; segment tips with hemispheric to globose osmophore cells arranged singly or in moniliform chains; labellum stiffly hinged to delicately balanced and tremulous, obscurely to distinctly three-lobed; lateral lobes broad, entire or with a few marginal teeth; midlobe often differently coloured to the base, with elongate or deltate marginal teeth subsimilar to the calli, decrescent to the apex, sometimes coalescent into a marginal band; calli separate, in rows, often extending onto the midlobe, stalked, clubbed or often gammate; basal calli often dissimilar to the rest; column with glandular hairs, broadly winged, the wings not extending above the anther, with a pair of prominent basal gland -like structures, usually yellow.
Size and Distribution
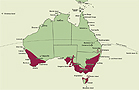
A genus of c. 200 species endemic to Australia and mainly distributed in southern areas. In eastern Australia the genus is distributed between about 28ş30' S on the Darling Downs in southern Queensland and 43ş30' S in southern Tasmania. In southwestern Western Australia the genus is distributed between about 27ş30' S near Kalbarri and 35ş S near Albany. The genus ranges from the coast to areas well inland and into the mountains. State occurrence: Queensland, New South Wales, Victoria, Tasmania, South Australia, Western Australia.
Ecology
These orchids grow in a range of forested and shrubby habitats, including open forest, woodland, heathy forest, mallee communities, Sheoak thickets, heathland and shrubland with some species occupying specific niches on stabilised coastal dunes, coastal headlands, in shallow soil over rock sheets and on granite outcrops. Most species grow in well-drained soils but some species, particularly in Western Australia, occur in soils that are wet to waterlogged in winter, the plants sometimes growing in standing water.
Biology
Pollination: Most species of Arachnorchis seem to be pollinated by the males of species of thynnine wasps in highly specific relationships with each orchid being adapted for pollination by a single species of wasp (Bower 2001). The male wasps respond to specific floral odours that mimic the sex pheromones given off by receptive female wasps (allomones). Females are wingless and when ready to mate climb on plants and emit sex pheromones to attract the males, one of which picks up the female and rapidly flies off with her, mating while in flight or later after landing. The males approach the receptive female flying into the wind in a zigzag pattern following the pheromone trail and they adopt the same pattern when approaching Arachnorchis flowers. The source of the floral odours in Arachnorchis are the hemispherical or globose glands concentrated along the tepal tips and in some species aggregated into specialised thickened club-like structures. When close to the flower visual stimuli including red labellum tips and clustered red calli mimic the female wasp and further deceive the male into attempting copulation with the labellum. In species with a relatively immobile labellum, such as Arachnorchis huegelii, the wasp crawls up the centre of the labellum using the rows of calli to gain purchase and moves towards the labellum base contacting the stigma and depositing any pollinia it carries. As it backs out a coating of glue from the rostellum is smeared onto its thorax just before it contacts the anther allowing deposition of pollinia (Stoutamire 1983). Another group of species, as exemplified by Arachnorchis tentaculata, have a delicately balanced labellum that tips forwards when a male wasp alights on the maroon apex. This causes the wasp to scramble forwards passing the point of balance of the labellum, thus tipping it back against the column and momentarily trapping the wasp. During its struggles to free itself pollinia can be picked up or deposited. Once the wasp has left the flower the labellum returns to its original position (Erickson 1965, Stoutamire 1983, Bower 1992).
It has been postulated that some species of Arachnorchis might be pollinated by pollen-feeding or nectar-feeding bees and flies (Stoutamire 1975, 1983), particularly large white-flowered species such as Arachnorchis longicauda. This needs further study in light of the high incidence of sexually deceptive species in the genus (Bower 2001). Two studies of the pollination of Arachnorchis rigida provided some evidence for a mixed syndrome involving both food deception (Bates 1984) and sexual deception (Bickerton 1997). At least one species of Arachnorchis, namely Arachnorchis graminifolia, is self-pollinating.
Reproduction: Reproduction is solely from seed. After pollination the capsule swells rapidly in an erect position and seed dispersal occurs 4-8 weeks later. There is no increase in the length of the peduncle or pedicel during this process. Apomixis is unknown in the genus.
Seasonal Growth: The prevailing climate where these orchids grow is strongly seasonal and the plants survive as dormant tubers from late spring or summer to autumn. The leaf emerges after autumn-winter rains and is fully developed by flowering time. The tubers are partly surrounded by a multi-layered fibrous tunic that is connected to the soil surface by a fibrous sheath. Each new tuber adds a layer of fibre to the inner side of the tunic and sheath.
Flowering: The majority of species flower in spring and early summer. One species, Arachnorchis drummondii, flowers in winter.
Hybrids: Natural hybrids are not uncommon between sympatric species of Arachnorchis, particularly in habitats that have been disturbed or degraded in some way (Heberle 1982). Many hybrids are rare and only occur sporadically, others are much more frequent and persist for several seasons. Backcrossing resulting in hybrid swarms is occasionally observed.
Fire: Many species flower freely in the absence of fire although a general enhancement is noticeable when densely overgrown sites are burnt. Sometimes these responses are dramatic and result in massed floral displays. A few species from Western Australia are strict fire-followers and flower only after a summer burn.
Derivation
Arachnorchis is derived from the Greek arachne, spider and Orchis, another genus of Orchidaceae and applied generally to an orchid.
Botanical Description
Perennial geophytic herbs, deciduous, sympodial. Plants hairy. Roots absent. Tubers globose, paired, fleshy, enclosed partially within a multi-layered persistent fibrous tunic; replacement tuber formed adjacent to the parent tuber within the upper part of the tunic on the end of a short dropper or below the parent tuber on a longer dropper which is fully enclosed by the fibrous sheath; daughter tubers absent. Stem erect, short, unbranched, with a membranous cataphyll at the apical node, covered by a multi-layered persistent fibrous tunic (sheath) to the soil surface, each new annual shoot extending through this sheath. Trichomes scattered to dense on most above-ground parts, mostly eglandular, glandular on the ovary, multicelled; basal cell enlarged, white, often ornamented. Flowering and non-flowering plants monomorphic. Sterile plants consist of a single leaf. Fertile plants consist of a basal leaf subtending an inflorescence. Leaf solitary, basal, sessile, green, often with purple basal blotches, rarely reddish abaxially. Leaf lamina mostly ovate, lanceolate or linear, subcoriaceous, convolute in bud; margins entire; trichomes eglandular, elongate, multicelled, with an enlarged, white, sculptured basal cell. Venation anastomosing. Inflorescence terminal, erect, racemose, 1-4-flowered. Peduncle wiry, hairy, the trichomes as on the leaf, with 1 spreading sterile bract. Floral bracts foliaceous, sheathing. Pedicel short, hairy. Ovary elongate, straight, ribbed, covered with glandular trichomes. Flowers resupinate, porrect, opening widely, brightly coloured (cream, white, green, pink, red, rarely yellowish), pedicellate. Sepals and petals subsimilar, elongate, broadest proximally then tapered and filiform, glandular distally, either extended into caudae or thickened into terminal club-like structures, both structures bearing hemispheric to globose osmophore cells singly or in moniliform chains. Dorsal sepal free, similar or subsimilar to the lateral sepals, erect, spreading or recurved, rarely drooping. Lateral sepals free, similar or subsimilar to the dorsal sepal, parallel or divergent, erect, porrect or drooping. Petals free, usually shorter and narrower than the sepals. Labellum free, attached by a short claw to the anterior column base, ranging from stiffly hinged to delicately balanced and tremulous in the slightest breeze, markedly dissimilar in size and shape to the sepals and petals, ecalcarate. Labellum lamina mostly ovate-cordate, obscurely lobed to strongly three-lobed; lateral lobes erect to spreading, sometimes distally thickened, entire or with short to long marginal teeth; midlobe usually recurved or incoiled, sometimes porrect, apex often maroon; marginal teeth linear, gammate or deltate, subsimilar to the calli, decrescent to the apex, sometimes coalescent into a marginal band. Callus of 2-several longitudinal rows of calli, often extending onto the midlobe; calli separate, stalked, clavate, gammate or distally curved, smooth or papillate, variously coloured (commonly maroon); basal 2-6 calli often different in shape, size and/or colour from the rest; distal calli decrescent, becoming sessile, irregular in size and arrangement. Nectar absent. Spur absent. Column lacking free filament and style, slender, incurved, mostly translucent with irregular markings. Column foot absent. Pseudospur absent. Column wings broad, not extending above the anther. Anther terminal, 4-celled, persistent, basifixed, erect, papillate, rostrate. Pollinarium absent. Pollinia 4, bilobed, flat, mealy, yellow. Viscidium absent. Stigma entire, circular, concave. Rostellum ventral. Capsules dehiscent, glandular-hairy, erect; peduncle not elongating in fruit; pedicel not elongating in fruit. Seeds numerous, light coloured, winged.
Taxonomy
The genus Arachnorchis was established as a segregate from Caladenia in 2001 (Joneset al. 2001). It is distinguished from Caladenia by the presence on the leaf and scape of multiseriate eglandular trichomes each with an enlarged white basal cell; large flowers with attenuate tepals in which the tips are extended into caudae or thickened into club-like units, both structures bearing specialised hemispheric to globose osmophore cells, sometimes arranged in moniliform chains; and, the column base with a pair of prominent gland -like structures, usually yellow. This change was based on the choice of Caladenia flava as the type of Caladenia (see also entry for Caladenia). This choice of the type for Caladenia and the narrow circumscription of the genus has been challenged by Hopper and Brown (2004).
Notes
A distinctive group within Arachnorchis, centred around Arachnorchis dilatata, has the labellum delicately balanced such that it is tremulous and can tremble or vibrate in a light breeze. The labellum is also distinctive in these species in that it is green with a maroon apex (sometimes separated by a white area) and the marginal teeth on the lateral lobes are long and narrow resembling the bars of a gaol.
Arachnorchis drummondii is unique within the genus for its winter-flowering habit and the leaf being only partially developed at anthesis. The leaf, the abaxial surface of which is heavily stained with anthocyanin pigments, continues growing and reaches its full size after flowering.
Nomenclature
Arachnorchis D.L.Jones and M.A.Clem., Orchadian 13(9): 392 (2001). Type species: Arachnorchis patersonii (R.Br.) D.L.Jones and M.A.Clem.
Phlebochilus (Lindl.) Szlach., Polish. Bot. J. 46(1): 14 (2001), pro parte.
Infrageneric Taxa: No infrageneric taxa are currently recognised.
References
Backhouse, G. and Jeanes, J. (1995). The Orchids of Victoria. Miegunyah Press, Carlton, Victoria.
Bates, R. (1984). The ecology and biology of Caladenia rigida (Orchidaceae). South Australian Naturalist 58: 56-65.
Bates, R.J. and Weber, J.Z. (1990). Orchids of South Australia. Government Printer, South Australia.
Bentham, G. (1873). Flora Australiensis, vol. 6. L.Reeve and Co, London.
Bickerton, D. (1997). A pollination study of the endangered orchid Caladenia rigida. Part 3: Pollinators. Journal of the Native Orchid Society of South Australia 21: 74-75.
Bishop, T. (1996). Field Guide to the Orchids of New South Wales and Victoria. University of New South Wales Press, Sydney.
Bower, C.C. (1992). The use of pollinators in the taxonomy of sexually deceptive orchids in the subtribe Caladeniinae (Orchidaceae). Orchadian 10: 331-338.
Bower, C.C. (2001). Pollination (Caladenia, pp. 93-97), in Pridgeon, A.M., Cribb, P.J., Chase, M.W. and Rasmussen, F.N. (eds), Genera Orchidacearum, vol. 2, Orchidoideae (part one). Oxford University Press, United Kingdom.
Clements, M.A. (1990). Catalogue of Australian Orchidaceae. Austral. Orch. Res. 1: 1-160.
Curtis, W.M. (1979). The Student’s Flora of Tasmania, Part 4A. Government Printer, Hobart.
Entwisle, T.J. (1994), in Walsh, N.G. and Entwisle, T.J. (1994). Flora of Victoria, vol. 2. Inkata Press, Melbourne.
Erickson, R. (1965). Orchids of the West. 2nd edn. Paterson Brokensha, Perth.
Heberle, R.L. (1982). Caladenia in WA and natural hybridisation. Orchadian 7: 78-83.
Hoffman, N. and Brown A.P. (1984, 1992). Orchids of South-west Australia. University of Western Australia Press, Perth.
Hopper, S.D. and Brown, A.P. (2004). Robert Brown's Caladenia revisited, including a revision of its sister genera Cyanicula, Ericksonella and Pheladenia (Caladeniinae: Orchidaceae). Aust. Syst. Bot. 17: 171-240.
Jones, D.L., Wapstra, H., Tonelli, P. and Harris, S. (1999). The Orchids of Tasmania. Melbourne University Press, Victoria.
Jones, D.L., Clements, M.A., Sharma, I.K. and Mackenzie, A.M. (2001). A new classification of Caladenia R.Br. (Orchidaceae). Orchadian 13(9): 389-419.
Lindley, J. (1830-40). The Genera and Species of Orchidaceous Plants. Ridgeways, London.
Nicholls, W.H. (1969). Orchids of Australia. Thomas Nelson, Melbourne.
Pfitzer, E.H.H. (1889). Orchidaceae. In A.Engler and K.Prantl (eds). Die Natürlichen Pflanzenfamilien. Leipzig.
Stoutamire, W.P. (1975). Pseudocopulation in Australian terrestrial orchids. American Orchid Society Bulletin 44: 226-233.
Stoutamire, W.P. (1983). Wasp-pollinated species of Caladenia in south-western Australia. Australian Journal of Botany 31: 383-394.
Weber, J.Z. and Bates, R. (1986), in Jessop, J.P. and Toelken, J.P. (eds), Flora of South Australia, 4th edn. South Australian Printing Division, Adelaide.
Willis, J.H. (1970). A Handbook to Plants in Victoria vol. 1 (2nd ed.). Melbourne University Press, Carlton.